

Pedunculopontine-stimulation obstructs hippocampal theta rhythm and halts movement
- PMID: 40410186
- PMCID: PMC12102179
- DOI: 10.1038/s41598-025-01695-8
Pedunculopontine-stimulation obstructs hippocampal theta rhythm and halts movement
Abstract
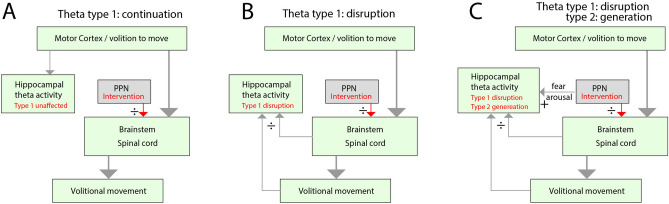
While the movement of rodents can be paused by optogenetic stimulation of a brainstem nucleus, the pedunculopontine nucleus (PPN), it is unknown whether this response has a functional purpose. The arrest appears conspicuously similar to fear-induced freezing behavior and could constitute a general halting mechanism for movement without an emotional component. Further, it is unclear to what extent brain activity is affected by the evoked motor arrest. Here, we investigate this phenomenon by engaging a distinct brain activity, the hippocampal theta rhythm. The theta rhythm is prominent during locomotor activity, absent under normal immobile situations, yet present under vigilant states like fear-induced freezing. Specifically, we ask whether the PPN-induced motor arrest has the same effect on the theta rhythm as if the animal would perform a volitional arrest, which results in the disappearance of the theta rhythm, or whether it would cause a continuation of the theta rhythm as would be expected by a fear-induced motor arrest. An alternative hypothesis is that the theta rhythm represents an ongoing intention to move rather than the movement itself. To distinguish between these two possibilities, we recorded the hippocampal brain rhythm before and during movement arrest induced by optogenetic stimulation of the PPN in rats. The PPN-induced motor arrest was associated with a clear obstruction of the ongoing theta activity. The timescale of movement arrest was less than 200 ms, similar to the obstruction of the theta rhythm. Since fear and behavioral freezing are associated with hippocampal theta rhythm, which we did not see during PPN stimulation, we suggest that induced motor arrest occurs without an associated emotional component. Further, our experiments reveal that the theta rhythm during motor activity does not represent an intention, but rather the ongoing sensory-motor state.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests. Declaration of AI Assistance in writing: Artificial intelligence (AI) in the form of generative Large Language Models (ChatGPT-4) assisted in writing this manuscript. The use of ChatGPT was done by rewriting a section for improving clarity. The output was reviewed and edited and adapted for meaning. All authors have read the manuscript and take full responsibility of the content.
Figures



Similar articles
-
Hippocampal theta rhythm induced by rostral pontine nucleus stimulation in the conditions of pedunculopontine tegmental nucleus inactivation.Brain Res Bull. 2013 Jul;96:10-8. doi: 10.1016/j.brainresbull.2013.04.005. Epub 2013 Apr 28. Brain Res Bull. 2013. PMID: 23632212
-
Modulation of hippocampal theta rhythm by the opioid system of the pedunculopontine tegmental nucleus.Acta Neurobiol Exp (Wars). 2007;67(4):447-60. doi: 10.55782/ane-2007-1661. Acta Neurobiol Exp (Wars). 2007. PMID: 18320722
-
Hippocampal theta rhythm after serotonergic activation of the pedunculopontine tegmental nucleus in anesthetized rats.Brain Res Bull. 2010 Oct 30;83(5):257-61. doi: 10.1016/j.brainresbull.2010.08.003. Epub 2010 Aug 10. Brain Res Bull. 2010. PMID: 20705121
-
Hippocampal formation theta activity and movement selection.Neurosci Biobehav Rev. 1998 Mar;22(2):221-31. doi: 10.1016/s0149-7634(97)00003-1. Neurosci Biobehav Rev. 1998. PMID: 9579313 Review.
-
What is the therapeutic mechanism of pedunculopontine nucleus stimulation in Parkinson's disease?Neurobiol Dis. 2019 Aug;128:67-74. doi: 10.1016/j.nbd.2018.06.014. Epub 2018 Jun 19. Neurobiol Dis. 2019. PMID: 29933055 Review.
Cited by
-
Cell-type specific sensory and motor activity in the cuneiform nucleus and pedunculopontine nucleus in mice.Sci Rep. 2025 Jul 1;15(1):20408. doi: 10.1038/s41598-025-05572-2. Sci Rep. 2025. PMID: 40594670 Free PMC article.
-
A Real-Time Vision-Based Adaptive Follow Treadmill for Animal Gait Analysis.Sensors (Basel). 2025 Jul 9;25(14):4289. doi: 10.3390/s25144289. Sensors (Basel). 2025. PMID: 40732417 Free PMC article.
References
-
- Josset, N. et al. Distinct contributions of mesencephalic locomotor region nuclei to locomotor control in the freely behaving mouse. Curr. Biol.28, 884–901. 10.1016/j.cub.2018.02.007 (2018). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
