

Multiple steps of dynein activation by Lis1 visualized by cryo-EM
- PMID: 40410592
- PMCID: PMC12350149
- DOI: 10.1038/s41594-025-01558-w
Multiple steps of dynein activation by Lis1 visualized by cryo-EM
Abstract
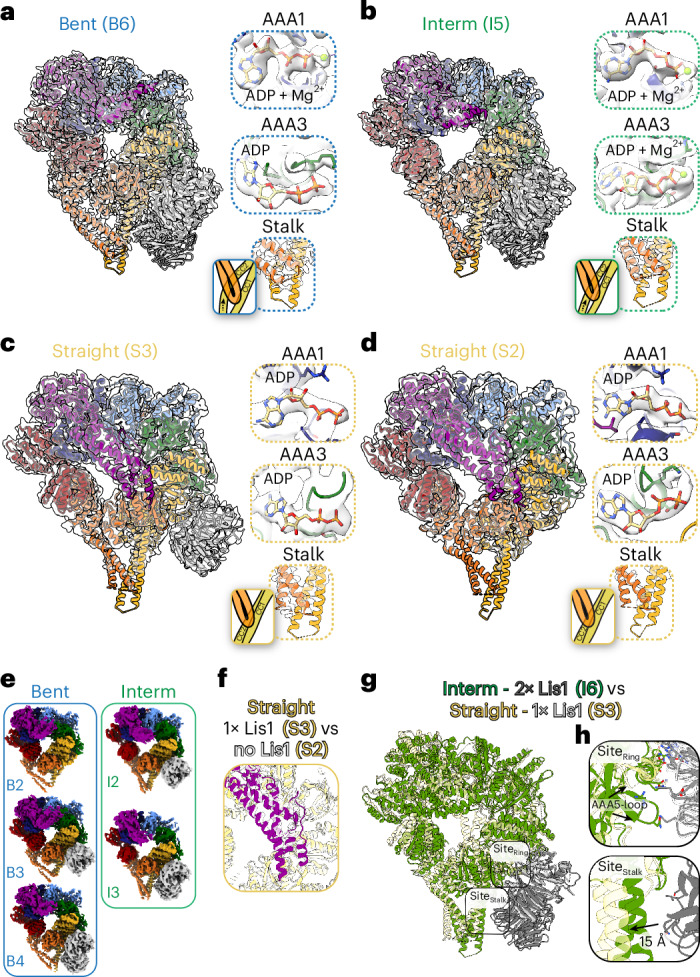
Cytoplasmic dynein-1 (dynein) is an essential molecular motor controlled in part by autoinhibition. Lis1, a key dynein regulator mutated in the neurodevelopmental disease lissencephaly, plays a role in dynein activation. We recently identified a structure of partially autoinhibited dynein bound to Lis1, which suggests an intermediate state in dynein's activation pathway. However, other structural information is needed to fully understand how Lis1 activates dynein. Here, we used cryo-EM and yeast dynein and Lis1 incubated with ATP at different time points to reveal conformations that we propose represent additional intermediate states in dynein's activation pathway. We solved 16 high-resolution structures, including 7 distinct dynein and dynein-Lis1 structures from the same sample. Our data support a model in which Lis1 relieves dynein autoinhibition by increasing its basal ATP hydrolysis rate and promoting conformations compatible with complex assembly and motility. Together, this analysis advances our understanding of dynein activation and the contribution of Lis1 to this process.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures












Update of
-
Cryo-EM visualizes multiple steps of dynein's activation pathway.bioRxiv [Preprint]. 2024 Oct 13:2024.09.28.615567. doi: 10.1101/2024.09.28.615567. bioRxiv. 2024. Update in: Nat Struct Mol Biol. 2025 Aug;32(8):1434-1444. doi: 10.1038/s41594-025-01558-w. PMID: 39416051 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
