

Single-cell profiling of bone metastasis ecosystems from multiple cancer types reveals convergent and divergent mechanisms of bone colonization
- PMID: 40412393
- PMCID: PMC12278635
- DOI: 10.1016/j.xgen.2025.100888
Single-cell profiling of bone metastasis ecosystems from multiple cancer types reveals convergent and divergent mechanisms of bone colonization
Abstract
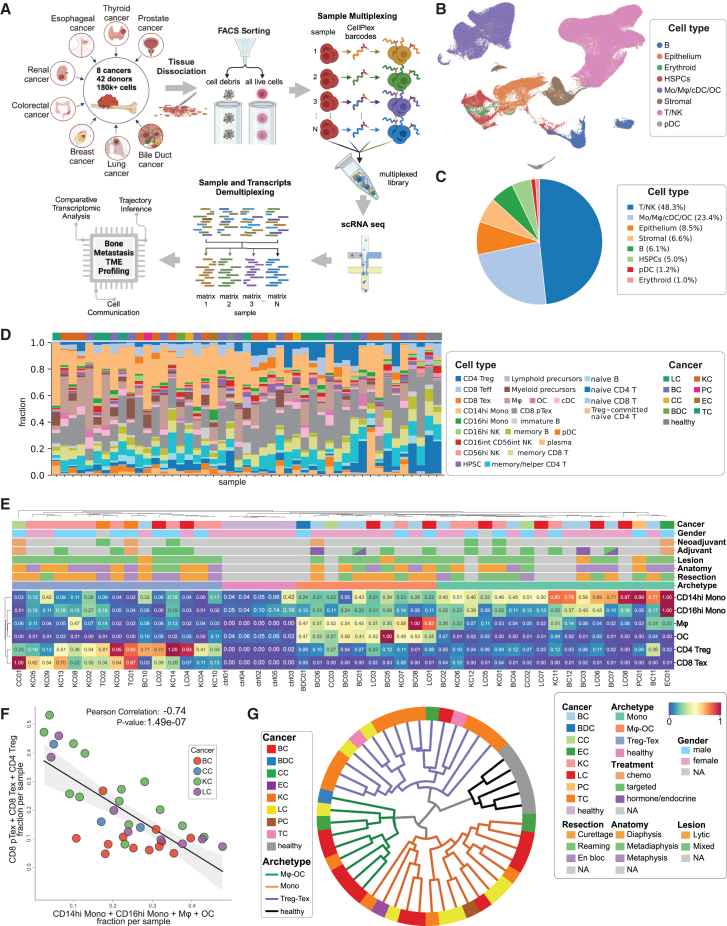
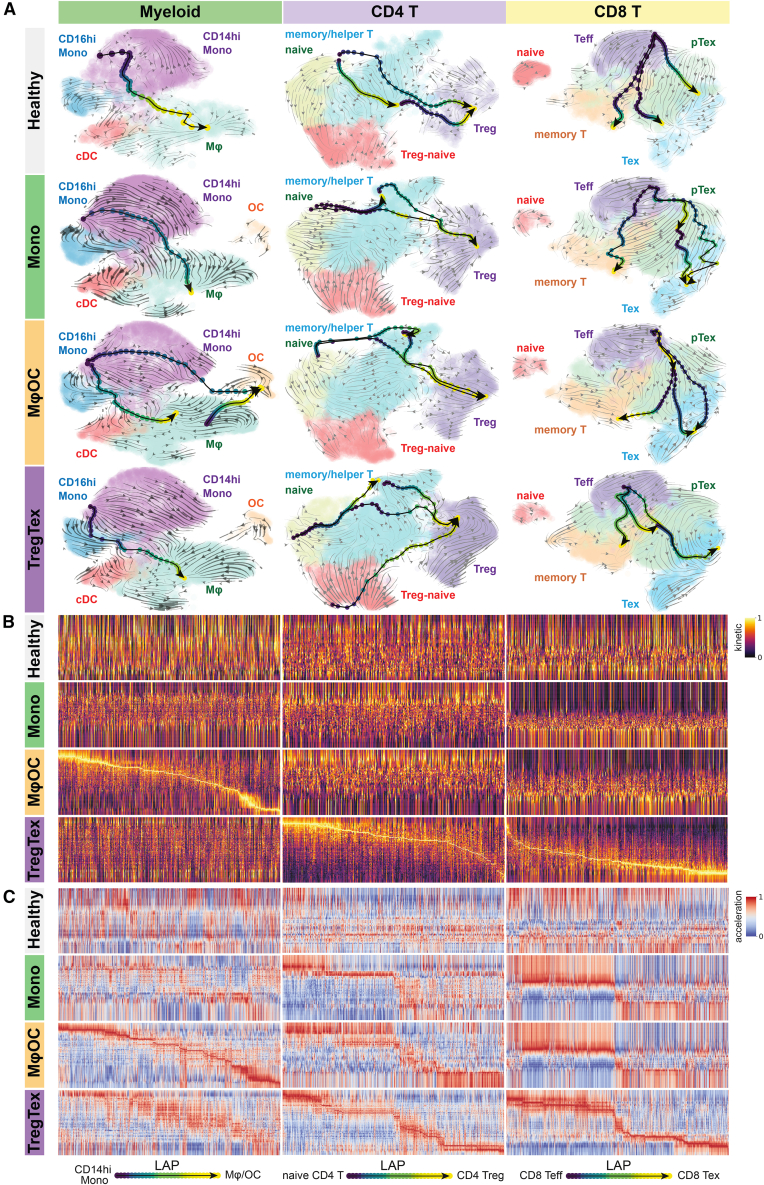
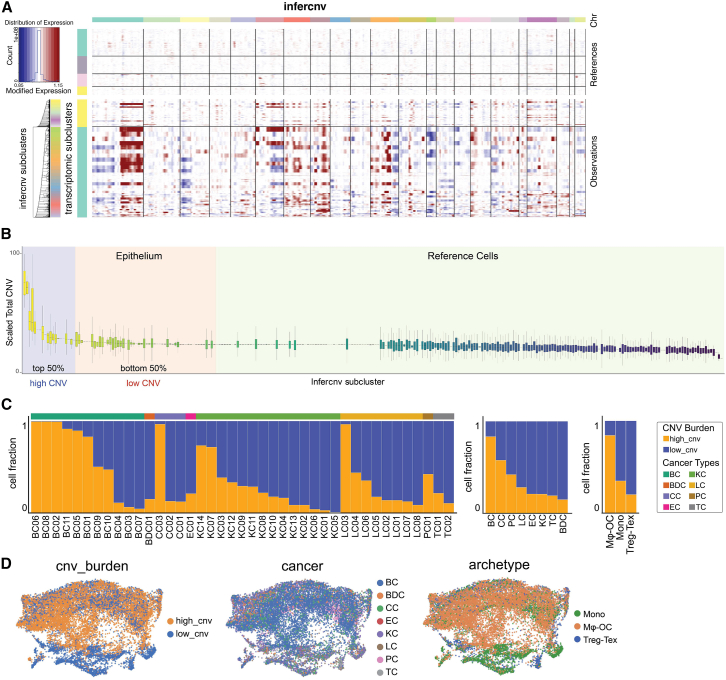
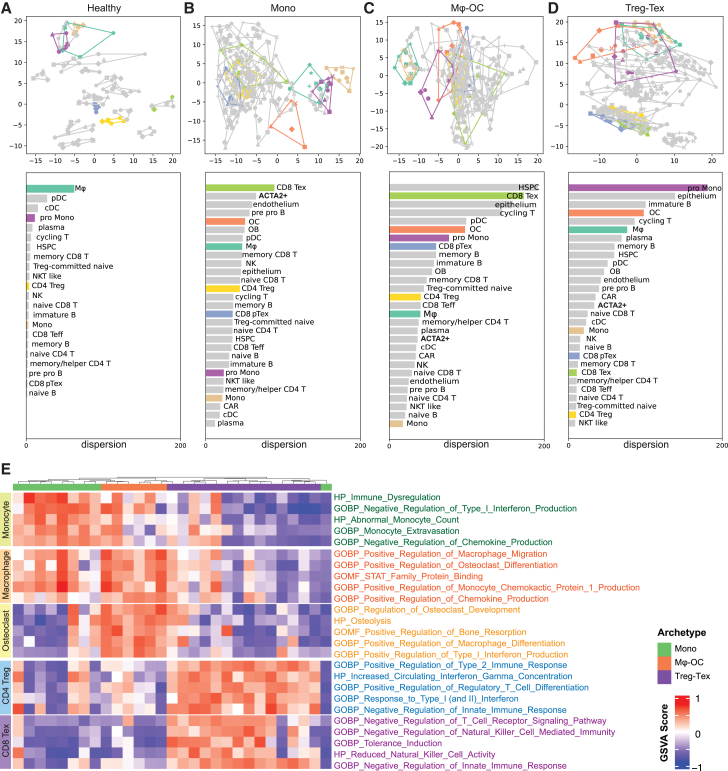
Bone is a common site for metastasis of solid cancers. The diversity of histological and molecular characteristics of bone metastases (BMs) remains poorly studied. Here, we performed single-cell RNA sequencing on 42 BMs from eight cancer types, identifying three distinct ecosystem archetypes, each characterized by an enrichment of specific immune cells: macrophages/osteoclasts, regulatory/exhausted T cells, or monocytes. We validated these archetypes by immunostaining on tissue sections and bioinformatic analysis of bulk RNA sequencing/microarray data from 158 BMs across more than 10 cancer types. Interestingly, we found only a modest correlation between the BM archetypes and the tissues of origin; BMs from the same cancer type often fell into different archetypes, while BMs from different cancer types sometimes converged on the same archetype. Additional analyses revealed parallel immunosuppression and bone remodeling mechanisms, some of which were experimentally validated. Overall, we discovered unappreciated heterogeneity of BMs across different cancers.
Keywords: bone metastases; convergent and divergent evolution; immune archetypes; immune evasion; immunosuppression; single-cell RNA sequencing; tumor microenvironment.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
