

Viral-mediated knockdown of Atxn2 attenuates TDP-43 pathology and muscle dysfunction in the PFN1C71G ALS mouse model
- PMID: 40413526
- PMCID: PMC12102926
- DOI: 10.1186/s40478-025-02005-z
Viral-mediated knockdown of Atxn2 attenuates TDP-43 pathology and muscle dysfunction in the PFN1C71G ALS mouse model
Abstract
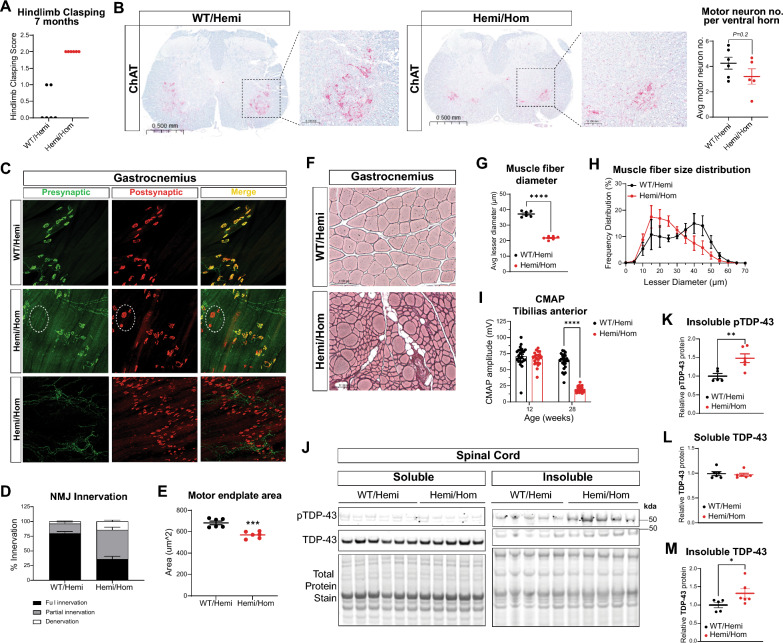
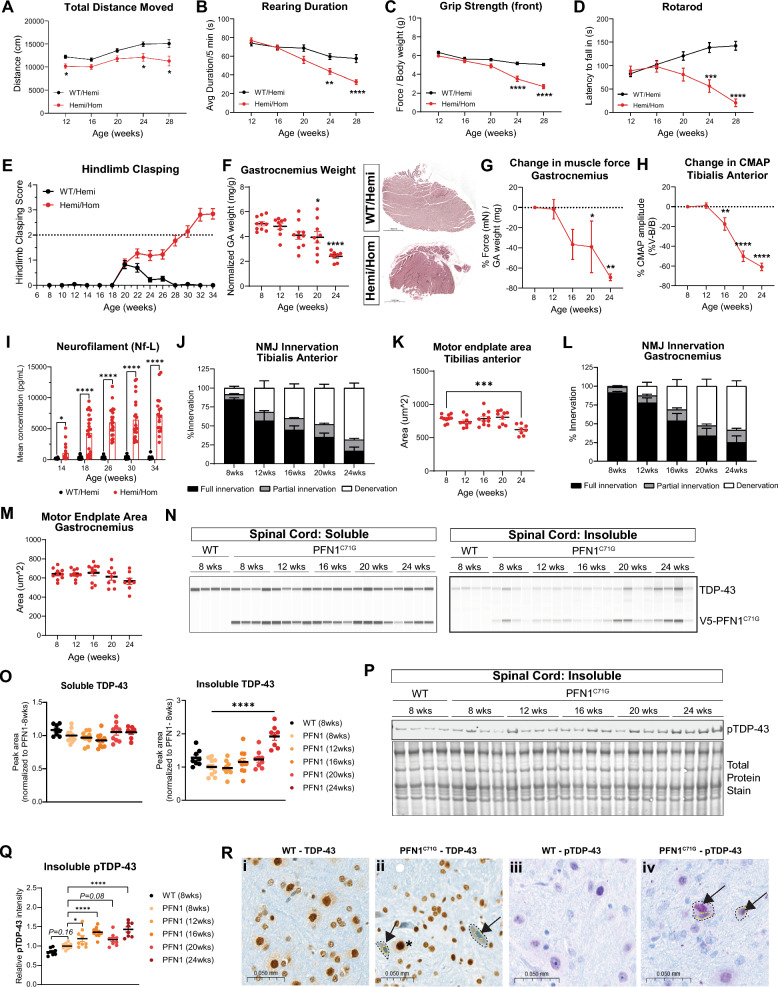
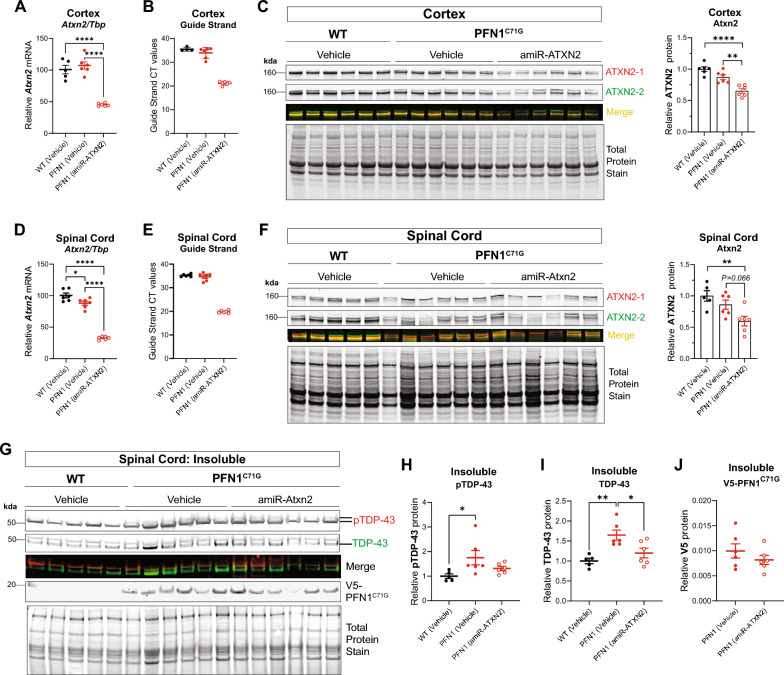
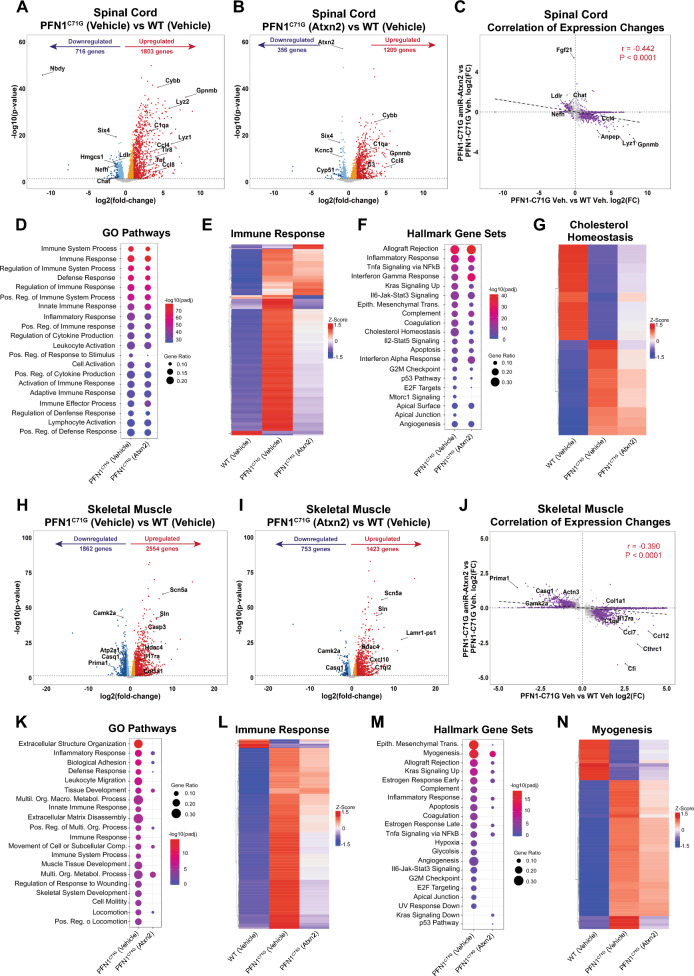
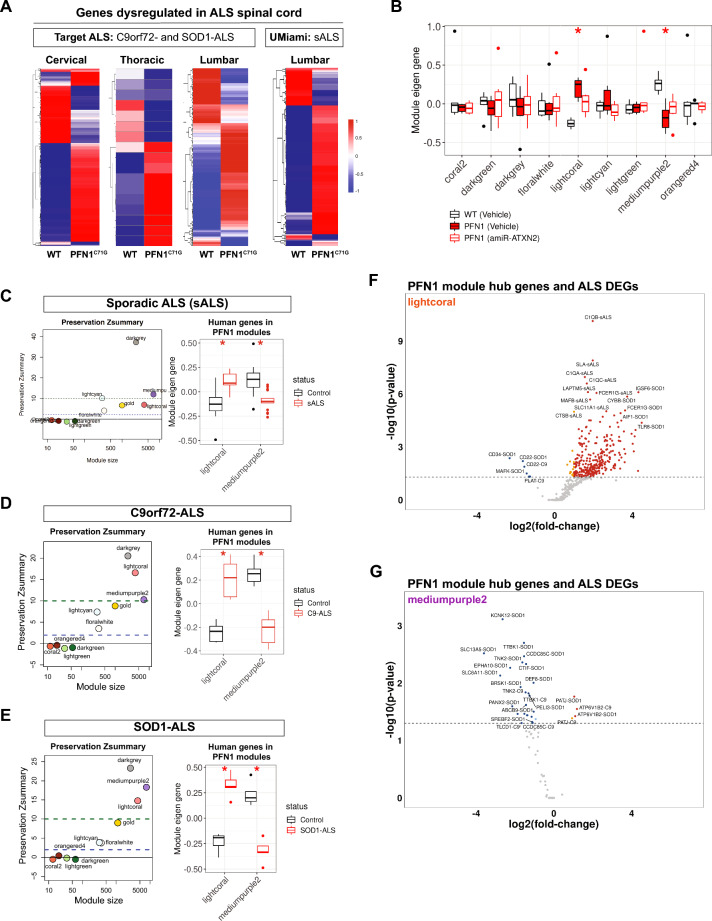
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disorder characterized by progressive motor neuron loss and muscle atrophy. Hyperphosphorylated aggregation of the RNA-binding protein, TDP-43, in the motor cortex and spinal cord are defining molecular features of ALS, suggesting TDP-43 dysfunction underlies disease pathogenesis. This phenomenon, however, has been difficult to recapitulate endogenously in animal models, impeding characterization of TDP-43 pathobiology in neurodegeneration. In this study, we report age-dependent accumulation of TDP-43 pathology in the spinal cord and progressive muscle-related deficits in transgenic mice expressing the ALS-associated PFN1C71G mutant protein. We show that transgenic neuronal expression of PFN1C71G induces early hyperphosphorylation of endogenous TDP-43 in the spinal cord that augments over time, preceding accumulation of insoluble non-phosphorylated TDP-43 and the manifestation of muscle denervation and motor dysfunction. Sustained knockdown of Atxn2 in the central nervous system (CNS) in pre-symptomatic PFN1C71G mice by AAV-driven expression of an artificial microRNA (AAV-amiR-Atxn2) reduces aberrant TDP-43 in the spinal cord, while delaying neurodegeneration and improving muscle and motor function. RNA-sequencing analysis of spinal cord samples from PFN1C71G mice and ALS donors show shared patterns of transcriptional perturbation, including a pro-inflammatory gene signature that is attenuated by AAV-amiR-Atxn2. Notably, impaired regulation of the PFN1C71G skeletal muscle transcriptome exceeds that of the spinal cord and is also improved by Atxn2 reduction in the CNS. Lastly, we find significant gene co-expression network homology between PFN1C71G mice and human ALS, with shared dysregulation of modules related to neuroinflammation and neuronal function and uncover novel hub genes that provide biological insight into ALS and potential drug targets that can be further investigated in this mouse model.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethical approval and consent to participate: All procedures were formed in accordance with NIH Guide for the Care and Use of Experimental Animals and studies were approved by the Biogen Institutional Animal Care and Use Committee (IACUC). Consent for publication: Not applicable. Competing interests: All authors are employees of Biogen (Cambridge, MA, USA).
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
