

At-RS31 orchestrates hierarchical cross-regulation of splicing factors and integrates alternative splicing with TOR-ABA pathways
- PMID: 40415535
- PMCID: PMC12177302
- DOI: 10.1111/nph.70221
At-RS31 orchestrates hierarchical cross-regulation of splicing factors and integrates alternative splicing with TOR-ABA pathways
Abstract
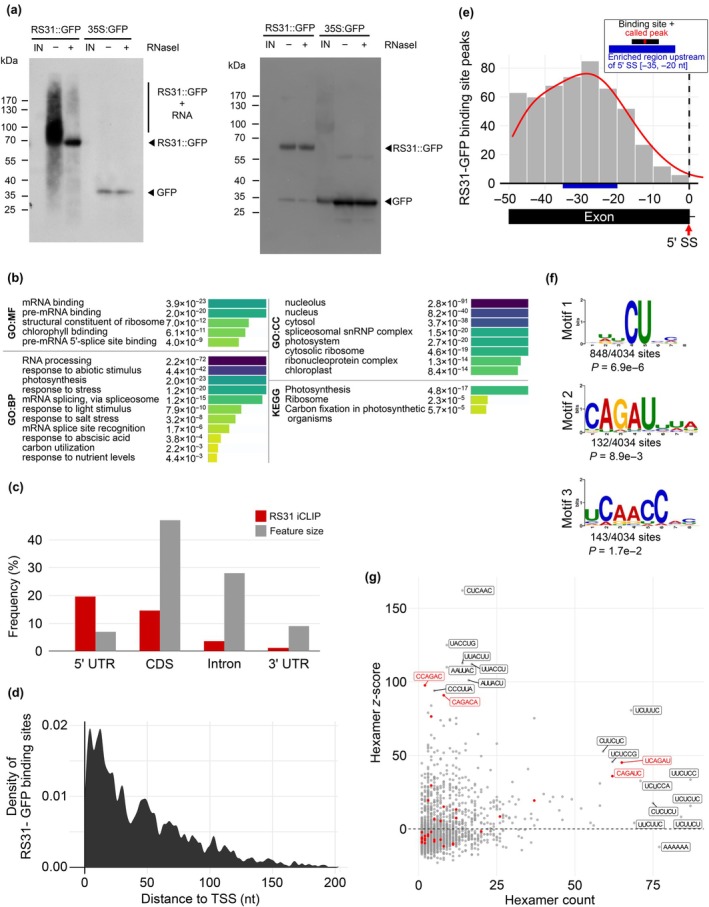
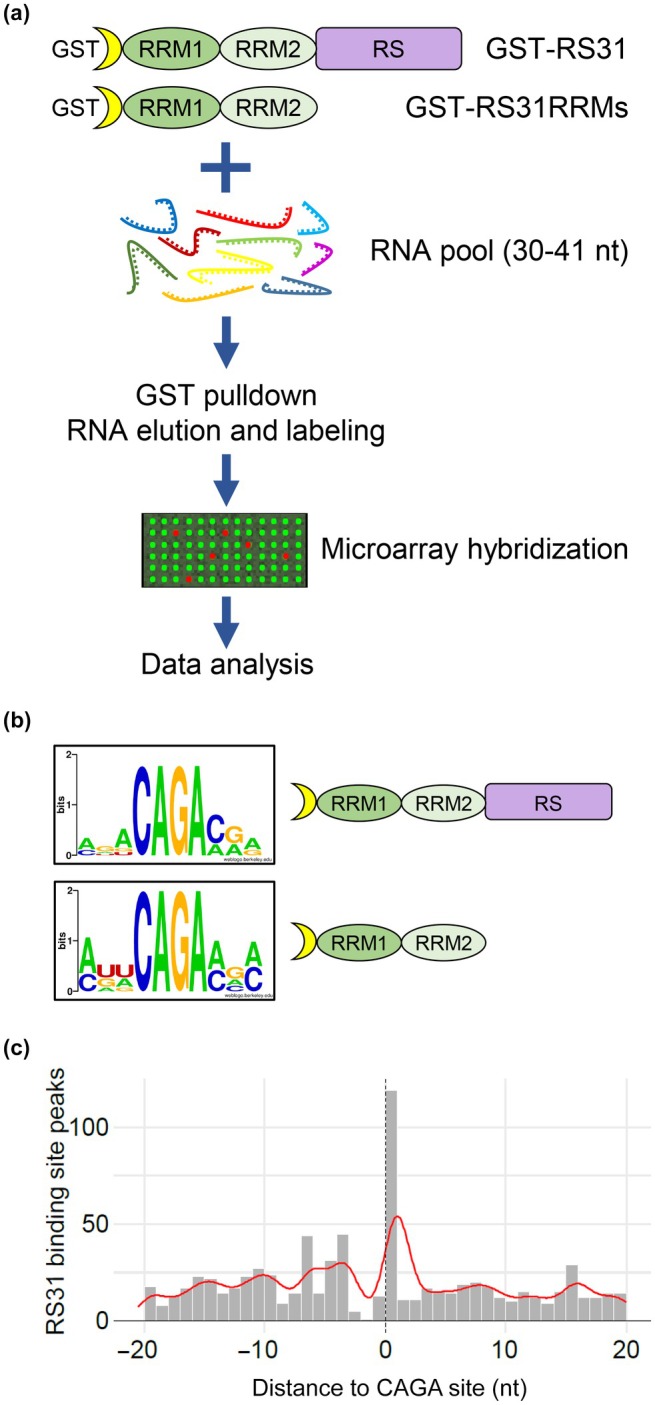
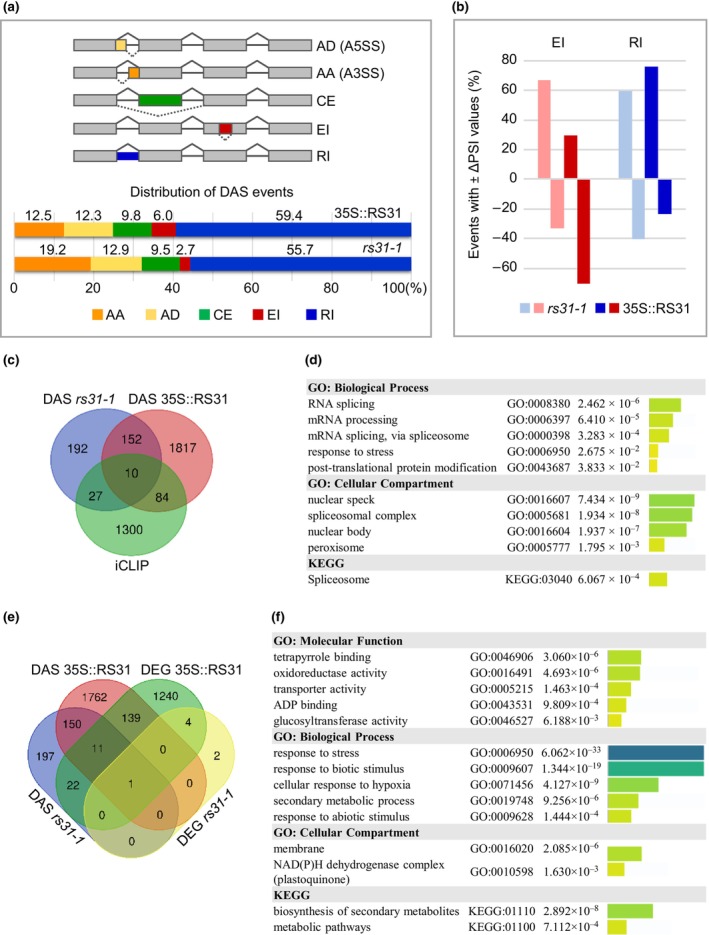
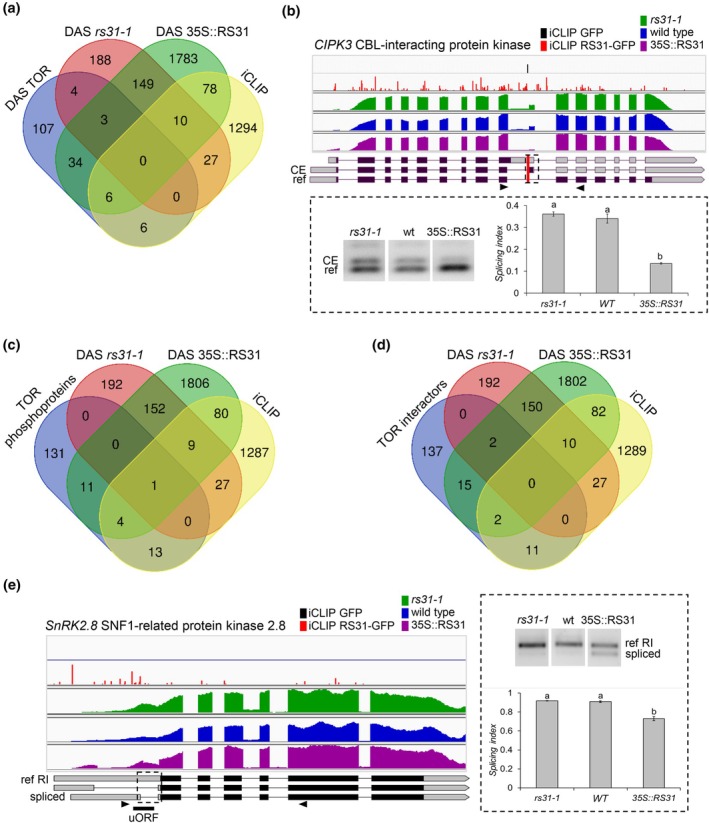
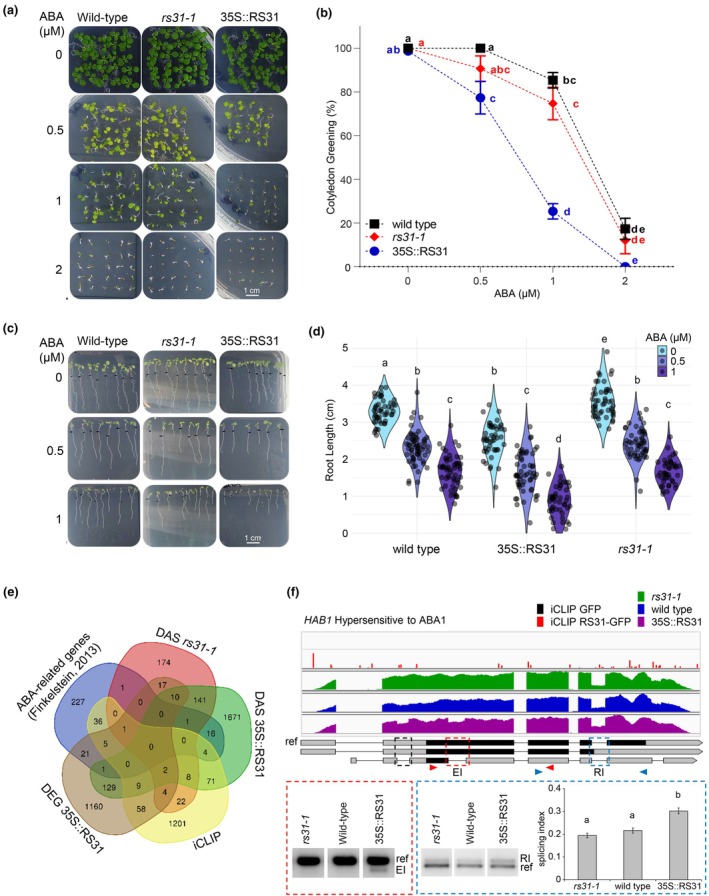
Alternative splicing is essential for plants, enabling a single gene to produce multiple transcript variants to boost functional diversity and fine-tune responses to environmental and developmental cues. Arabidopsis thaliana At-RS31, a plant-specific splicing factor in the Serine/Arginine-rich (SR) protein family, responds to light and the Target of Rapamycin (TOR) signalling pathway, yet its downstream targets and regulatory impact remain unknown. To identify At-RS31 targets, we applied individual-nucleotide resolution crosslinking and immunoprecipitation (iCLIP) and RNAcompete assays. Transcriptomic analyses of At-RS31 mutant and overexpressing plants further revealed its effects on alternative splicing. iCLIP identified 4034 At-RS31 binding sites across 1421 genes, enriched in CU-rich and CAGA RNA motifs. Comparative iCLIP and RNAcompete data indicate that the arginine/serine (RS) domain of At-RS31 may influence its binding specificity in planta, underscoring the value of combining in vivo and in vitro approaches. Transcriptomic analysis showed that At-RS31 modulates diverse splicing events, particularly intron retention and exitron splicing, and influences other splicing modulators, acting as a hierarchical regulator. By regulating stress response genes and genes in both TOR and abscisic acid signalling pathways, At-RS31 may help integrate these signals, balancing plant growth with environmental adaptability through alternative splicing.
Keywords: ABA; Arabidopsis thaliana; RNAcompete; SR proteins; TOR kinase; alternative splicing; binding site; iCLIP.
© 2025 The Author(s). New Phytologist © 2025 New Phytologist Foundation.
Conflict of interest statement
None declared.
Figures

Update of
-
At-RS31 orchestrates hierarchical cross-regulation of splicing factors and integrates alternative splicing with TOR-ABA pathways.bioRxiv [Preprint]. 2024 Dec 7:2024.12.04.626797. doi: 10.1101/2024.12.04.626797. bioRxiv. 2024. Update in: New Phytol. 2025 Jul;247(2):738-759. doi: 10.1111/nph.70221. PMID: 39677721 Free PMC article. Updated. Preprint.
References
-
- Altmann M, Altmann S, Rodriguez PA, Weller B, Elorduy Vergara L, Palme J, Marin‐de la Rosa N, Sauer M, Wenig M, Villaecija‐Aguilar JA et al. 2020. Extensive signal integration by the phytohormone protein network. Nature 583: 271–276. - PubMed
-
- Barta A, Kalyna M, Lorkovic ZJ. 2008. Plant SR proteins and their functions. Current Topics in Microbiology and Immunology 326: 83–102. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
