

Regulation of TFEB in human placental Cytotrophoblasts and Syncytiotrophoblasts
- PMID: 40415650
- PMCID: PMC12104664
- DOI: 10.14814/phy2.70383
Regulation of TFEB in human placental Cytotrophoblasts and Syncytiotrophoblasts
Abstract
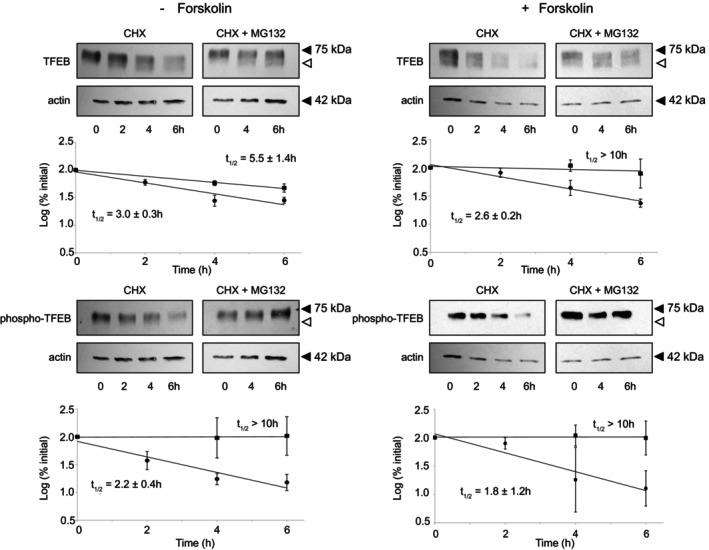
While cellular proteins exist in a dynamic state maintained by the balance of synthesis and degradation, there is a paucity of information on these processes in placental trophoblasts, including within cytotrophoblasts which differentiate into multi-nucleate syncytiotrophoblasts. TFEB, a transcription factor with a myriad of cellular activities, is one of the most abundant genes expressed in syncytiotrophoblasts compared to cytotrophoblasts. TFEB is localized to the nucleus of human BeWo differentiated syncytiotrophoblasts and to the cytoplasm of the undifferentiated cytotrophoblasts. Within both the cytotrophoblasts and syncytiotrophoblasts, TFEB exists in subcellular compartments as both phosphorylated and unphosphorylated forms and translocates between cytoplasm and nucleus upon amino acid starvation/refeeding. Endogenous TFEB and endogenous phospho-TFEB are both rapidly (t1/2 ~ 2-3 h) degraded via the ubiquitin proteasome system in cytotrophoblasts and in syncytiotrophoblasts. These results suggest dynamic regulatory processes during trophoblast development/differentiation.
Keywords: cytotrophoblast; degradation; placental; syncytiotrophoblast; ubiquitin.
© 2025 The Author(s). Physiological Reports published by Wiley Periodicals LLC on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





References
-
- Azar, C. , Valentine, M. , Trausch‐Azar, J. , Druley, T. , Nelson, D. M. , & Schwartz, A. L. (2018). RNA‐Seq identifies genes whose proteins are transformative in the differentiation of cytotrophoblast to syncytiotrophoblast, in human primary villous and BeWo trophoblasts. Scientific Reports, 8(1), 5142. 10.1038/S41598-018-23379-2 - DOI - PMC - PubMed
-
- Azar, C. , Valentine, M. C. , Trausch‐Azar, J. , Rois, L. , Mahjoub, M. , Nelson, D. M. , & Schwartz, A. L. (2021). RNA‐Seq identifies genes whose proteins are upregulated during syncytia development in murine C2C12 myoblasts and human BeWo trophoblasts. Physiological Reports, 9(1), e14671. 10.14814/phy2.14671 - DOI - PMC - PubMed
-
- Breitschopf, K. , Bengal, E. , Ziv, T. , Admon, A. , & Ciechanover, A. (1998). A novel site for ubiquitination: The N‐terminal residue, and not internal lysines of MyoD, is essential for conjugation and degradation of the protein. EMBO Journal, 17(20), 5964–5973. 10.1093/emboj/17.20.5964 - DOI - PMC - PubMed
-
- Cesana, M. , Tufano, G. , Panariello, F. , Zampelli, N. , Soldati, C. , Mutarelli, M. , Montefusco, S. , Grieco, G. , Sepe, L. V. , Rossi, B. , Nusco, E. , Rossignoli, G. , Panebianco, G. , Merciai, F. , Salviati, E. , Sommella, E. M. , Campiglia, P. , Martello, G. , Cacchiarelli, D. , … Ballabio, A. (2024). TFEB controls syncytiotrophoblast formation and hormone production in placenta. Cell Death and Differentiation, 31(11), 1439–1451. 10.1038/s41418-024-01337-y - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
