

Evolutionary insights into the selectivity of sterol oxidising cytochrome P450 enzymes based on ancestral sequence reconstruction
- PMID: 40417289
- PMCID: PMC12100521
- DOI: 10.1039/d5sc01863c
Evolutionary insights into the selectivity of sterol oxidising cytochrome P450 enzymes based on ancestral sequence reconstruction
Abstract
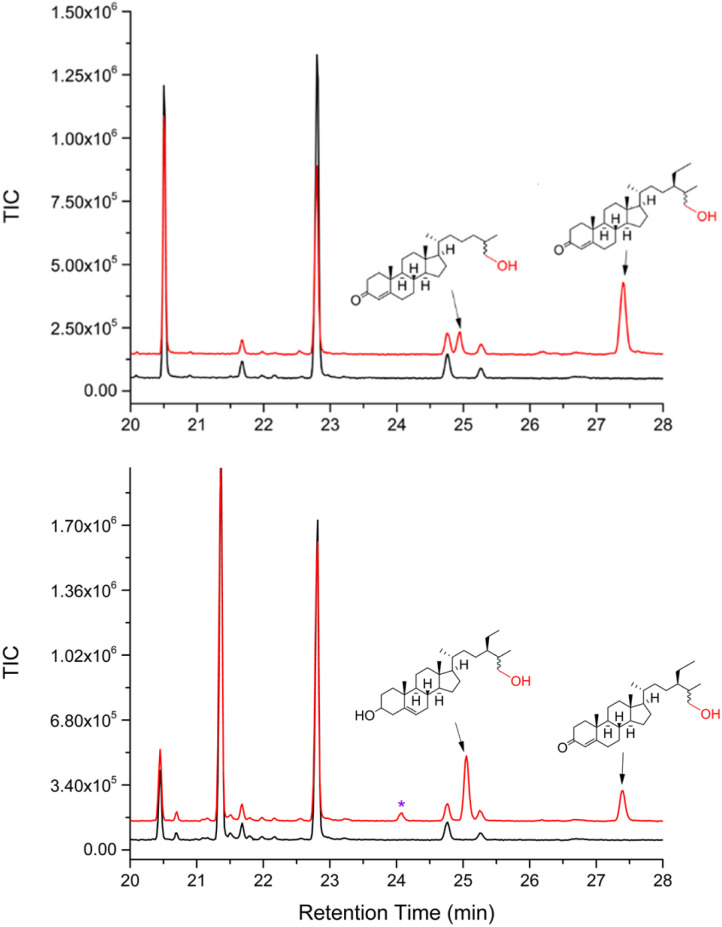
The cytochrome P450 (CYP) enzyme CYP125A1 is a crucial enzyme for the long-term survival and pathogenicity of Mycobacterium tuberculosis. CYP125 genes are found not only in pathogenic mycobacteria but are also widely dispersed within the Actinobacteria phylum, with many species possessing multiple copies of CYP125 encoding genes. Their primary function is the catalytic hydroxylation of the terminal methyl group of cholesterol and phytosterols. We have previously shown that CYP125 enzymes from distinct mycobacteria have substrate selectivity preferences for animal versus plant steroid oxidation. An evolutionary understanding of this selectivity is not known. Here, we use Ancestral Sequence Reconstruction (ASR), to support the hypothesis that some CYP125 enzymes evolved in a manner reflective of their adaptation to a pathogenic niche. We constructed a maximum-likelihood, most-recent common ancestor of the CYP125 clade (CYP125MRCA). We were then able to produce and characterise this enzyme both functionally and structurally. We found that CYP125MRCA was able to catalyse the terminal hydroxylation of cholesterol, phytosterols, and vitamin D3 (cholecalciferol); the latter was hydroxylated at both C-25 and C-26. This is the first example to date of vitamin D3 oxidation by a CYP125 enzyme, thereby demonstrating an increased substrate range of CYP125MRCA relative to its characterised extant relatives. The X-ray crystal structures of CYP125MRCA bound with sitosterol and vitamin D3 were determined, providing important insight into the changes that enable the expanded substrate range.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts to declare.
Figures




References
-
- McLeod M. P. Warren R. L. Hsiao W. W. Araki N. Myhre M. Fernandes C. Miyazawa D. Wong W. Lillquist A. L. Wang D. Dosanjh M. Hara H. Petrescu A. Morin R. D. Yang G. Stott J. M. Schein J. E. Shin H. Smailus D. Siddiqui A. S. Marra M. A. Jones S. J. Holt R. Brinkman F. S. Miyauchi K. Fukuda M. Davies J. E. Mohn W. W. Eltis L. D. Proc. Natl. Acad. Sci. U. S. A. 2006;103:15582–15587. doi: 10.1073/pnas.0607048103. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources