

Alterations of myocardial ketone metabolism in heart failure with preserved ejection fraction (HFpEF)
- PMID: 40420397
- PMCID: PMC12287784
- DOI: 10.1002/ehf2.15319
Alterations of myocardial ketone metabolism in heart failure with preserved ejection fraction (HFpEF)
Abstract
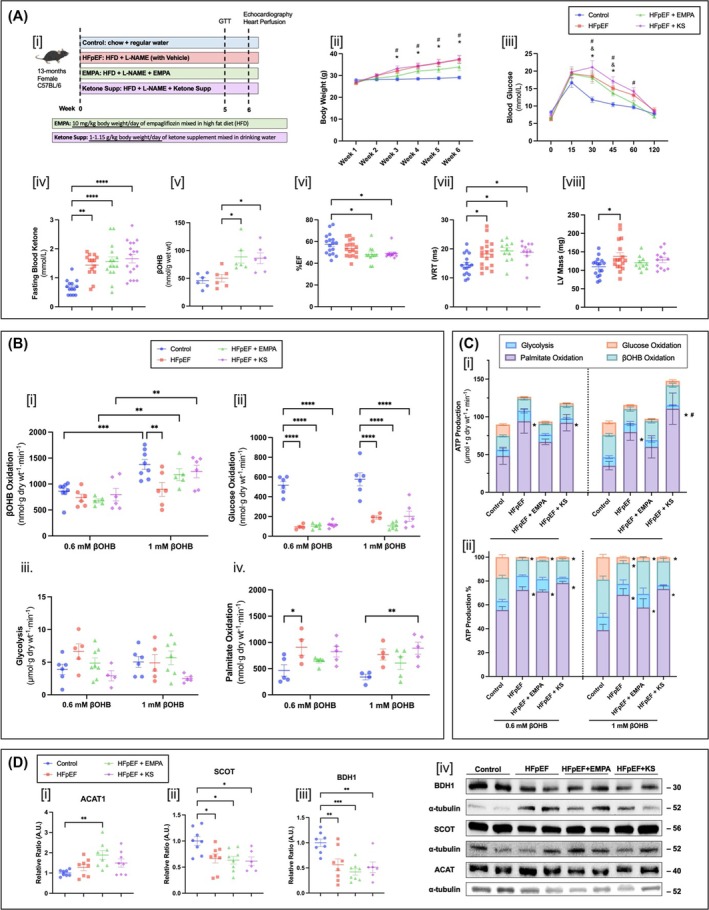
Introduction: Cardiac energy metabolism is disrupted in heart failure with preserved ejection fraction (HFpEF), as characterized by a switch from glucose oxidation towards fatty acid oxidation. However, although oxidation of ketones is an important source of ATP it remains unclear how the heart oxidizes ketones in HFpEF. It is also unclear whether elevating ketone supply to the heart can improve cardiac energetics and/or provide functional benefit for the hearts in HFpEF.
Aims: The present study investigated the effects of increasing ketone supply to the heart via ketone supplementation or SGLT2 inhibitor treatment in a mouse model of HFpEF.
Methods: HFpEF was induced in 13-month-old C57BL/6N female mice with 60% high-fat diet and L-NAME (0.5 g/L/day in the drinking water) for 6 weeks. In parallel, two other groups of mice were maintained on the HFpEF protocol while also receiving either a ketone ester supplement (1-3 butanediol 1 g/kg/day) or SGLT2 inhibitor (empagliflozin 10 mg/kg/day) for 6 weeks. Control mice were fed with regular low-fat diet and regular drinking water. Hearts of the mice were excised and perfused in the isolated working mode aerobically with 5-mM glucose, 0.8-mM palmitate, 100-μU/mL insulin, with either low (0.6 mM) or high (1 mM) levels of β-hydroxybutyrate. Metabolic rates of the hearts were measured with radiolabelled [U-14C] glucose, [9,10-3H] palmitate and [3-14C] β-hydroxybutyrate.
Results: In HFpEF mouse hearts, glucose oxidation was significantly decreased with a parallel increase in fatty acid oxidation. Increasing β-hydroxybutyrate levels from 0.6 to 1 mM in the perfusate resulted in a rise in ketone oxidation rates in control hearts (from 861 ± 63 to 1377 ± 94 nmol g dry wt-1 min-1), which was muted in HFpEF hearts (from 737 ± 68 to 897 ± 134 nmol g dry wt-1 min-1). Following ketone ester supplement or SGLT2 inhibitor treatment, HFpEF mice presented with restored ketone oxidation rates (from 674 ± 36 to 1181 ± 115 nmol g dry wt-1 min-1 with ketone ester supplement and from 797 ± 121 to 1240 ± 120 nmol g dry wt-1 min-1 with SGLT2i). Yet, this was not associated with improvement in cardiac function.
Conclusions: In HFpEF mice, the heart switches from glucose oxidation to fatty acid oxidation, with ketone oxidation being impaired. Increasing ketone supply to the heart via ketone ester supplementation or SGLT2 inhibitor treatment increases myocardial ketone oxidation rates but was not associated with functional improvements. Unlike HFrEF, ketone supplementation strategies may be less effective in HFpEF due to an impairment of myocardial ketone oxidation in HFpEF.
© 2025 The Author(s). ESC Heart Failure published by John Wiley & Sons Ltd on behalf of European Society of Cardiology.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous