

Lipid exposure leads to metabolic dysfunction in fetal sheep cardiomyocytes
- PMID: 40420618
- PMCID: PMC12106950
- DOI: 10.14814/phy2.70386
Lipid exposure leads to metabolic dysfunction in fetal sheep cardiomyocytes
Abstract
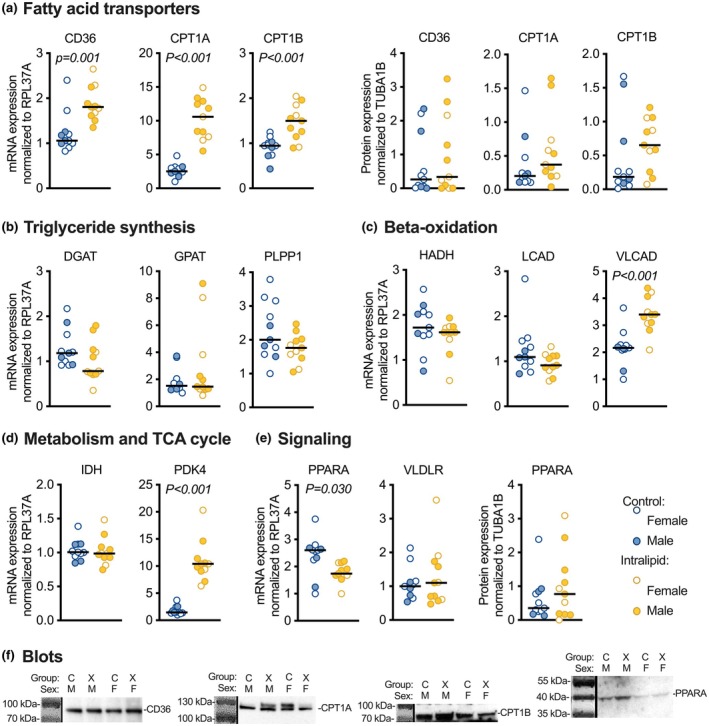
Fetal circulating lipids are low but rise precipitously following birth. It is unknown how prematurely elevated lipids affect the fetal heart, which primarily uses carbohydrates for energy. Fetal sheep were surgically instrumented and received Intralipid 20® or Lactated Ringer's Solution intravenously. After 8 days, myocardial biopsies were taken, and cardiomyocytes were dispersed. Lipid uptake was assessed by labeled saturated long-chain fatty acids (LCFA) and very long-chain fatty acids (VLCFA) incorporation. Maximal oxygen consumption rates (OCR) were measured. Gene and protein expression levels were measured by quantitative PCR and Western blotting. Intralipid treatment increased LCFA (p < 0.001) and VLCFA (p < 0.001) lipid droplet number, and LCFA (males p = 0.002) and VLCFA (p = 0.018) droplet size. Fetal Intralipid treatment reduced maximal OCR in basal media (p = 0.005). Palmitic acid decreased maximal OCR regardless of fetal treatment or length of in vitro exposure (p = 0.006). Fetal Intralipid upregulated genes included CD36 (p = 0.001), CPT1A (p < 0.001), CPT1B (p < 0.001), VLCAD (p < 0.001), and PDK4 (p < 0.001), with no differences in protein expression. There were no effects on ER stress, apoptosis, or autophagy markers. Extended elevated lipid levels in the fetus increased lipid uptake and may have shifted substrate preference towards lipids, but all lipid exposure depressed fetal cardiac metabolism. Prematurely elevated lipids mature but suppress oxidative metabolism.
Keywords: cardiomyocyte; fetus; metabolism; parenteral nutrition.
© 2025 The Author(s). Physiological Reports published by Wiley Periodicals LLC on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Myocardial Growth Response to Fetal Intralipid Infusion in Sheep.FASEB J. 2025 Aug 15;39(15):e70911. doi: 10.1096/fj.202501607R. FASEB J. 2025. PMID: 40879101
-
Thyroid hormone increases fatty acid use in fetal ovine cardiac myocytes.Physiol Rep. 2023 Nov;11(22):e15865. doi: 10.14814/phy2.15865. Physiol Rep. 2023. PMID: 38010207 Free PMC article.
-
Maturation of lipid metabolism in the fetal and newborn sheep heart.Am J Physiol Regul Integr Comp Physiol. 2023 Dec 1;325(6):R809-R819. doi: 10.1152/ajpregu.00122.2023. Epub 2023 Oct 23. Am J Physiol Regul Integr Comp Physiol. 2023. PMID: 37867472 Free PMC article.
-
Impact of new-generation lipid emulsions on cellular mechanisms of parenteral nutrition-associated liver disease.Adv Nutr. 2014 Jan 1;5(1):82-91. doi: 10.3945/an.113.004796. Adv Nutr. 2014. PMID: 24425726 Free PMC article. Review.
-
Use of Lipids in Neonates Requiring Parenteral Nutrition.JPEN J Parenter Enteral Nutr. 2020 Feb;44 Suppl 1:S45-S54. doi: 10.1002/jpen.1759. JPEN J Parenter Enteral Nutr. 2020. PMID: 32049399 Review.
References
-
- Bartelds, B. , Gratama, J. W. , Knoester, H. , Takens, J. , Smid, G. B. , Aarnoudse, J. G. , Heymans, H. S. , & Kuipers, J. R. (1998). Perinatal changes in myocardial supply and flux of fatty acids, carbohydrates, and ketone bodies in lambs. The American Journal of Physiology, 274, H1962–H1969. - PubMed
-
- Bartelds, B. , Knoester, H. , Smid, G. B. , Takens, J. , Visser, G. H. , Penninga, L. , van der Leij, F. R. , Beaufort‐Krol, G. C. , Zijlstra, W. G. , Heymans, H. S. , & Kuipers, J. R. (2000). Perinatal changes in myocardial metabolism in lambs. Circulation, 102(8), 926–931. 10.1161/01.cir.102.8.926 - DOI - PubMed
-
- Bosma, M. , Dapito, D. H. , Drosatos‐Tampakaki, Z. , Huiping‐Son, N. , Huang, L. S. , Kersten, S. , Drosatos, K. , & Goldberg, I. J. (2014). Sequestration of fatty acids in triglycerides prevents endoplasmic reticulum stress in an in vitro model of cardiomyocyte lipotoxicity. Biochimica et Biophysica Acta, 1841(12), 1648–1655. 10.1016/j.bbalip.2014.09.012 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
