

A conserved nuclease facilitates environmental DNA uptake
- PMID: 40421801
- PMCID: PMC12107432
- DOI: 10.1093/nar/gkaf443
A conserved nuclease facilitates environmental DNA uptake
Abstract
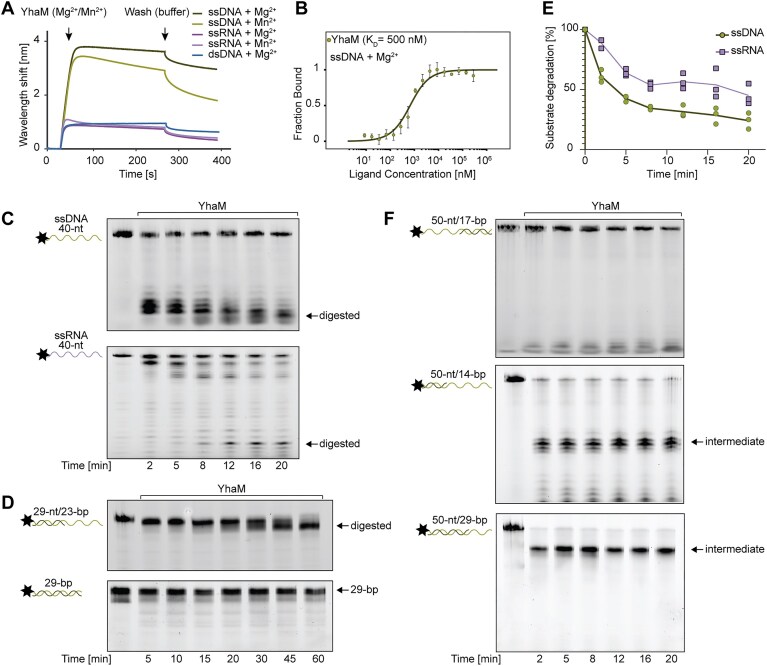
Bacteria acquire new traits through the uptake of genetic material from the environment, a process requiring DNA processing. However, the molecular inventory mediating this process is far from being completely understood. Here, we identify YhaM in Bacillus subtilis as a conserved 3'-deoxyribonuclease essential for the uptake and processing of genetic information in the form of single-stranded DNA. Our results show that YhaM assembles into hexamers in the presence of divalent cations, enhancing substrate binding, which is achieved through its conserved oligonucleotide-binding domain. Cells lacking YhaM show a severe defect in the uptake of plasmids and genomic DNA, but the transduction of double-stranded DNA by the phage SPP1 remains unaffected. These findings highlight a critical role of YhaM in single-stranded DNA maturation during natural transformation. Importantly, this function is conserved in various Gram-positive human pathogens such as Staphylococcus aureus, suggesting that it could contribute to the spread of antibiotic resistance.
© The Author(s) 2025. Published by Oxford University Press on behalf of Nucleic Acids Research.
Conflict of interest statement
None declared.
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
