

A Two-State Random Walk Model of Sperm Search on Confined Domains
- PMID: 40422493
- PMCID: PMC12110710
- DOI: 10.3390/e27050539
A Two-State Random Walk Model of Sperm Search on Confined Domains
Abstract
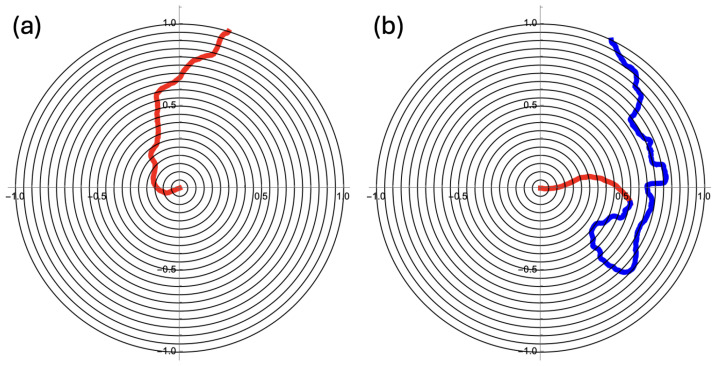
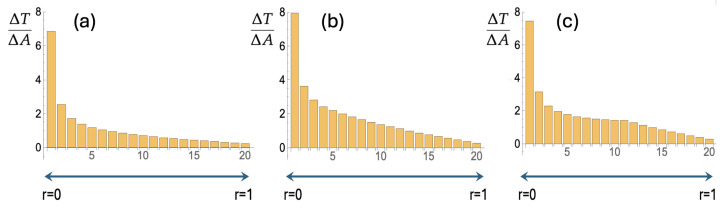
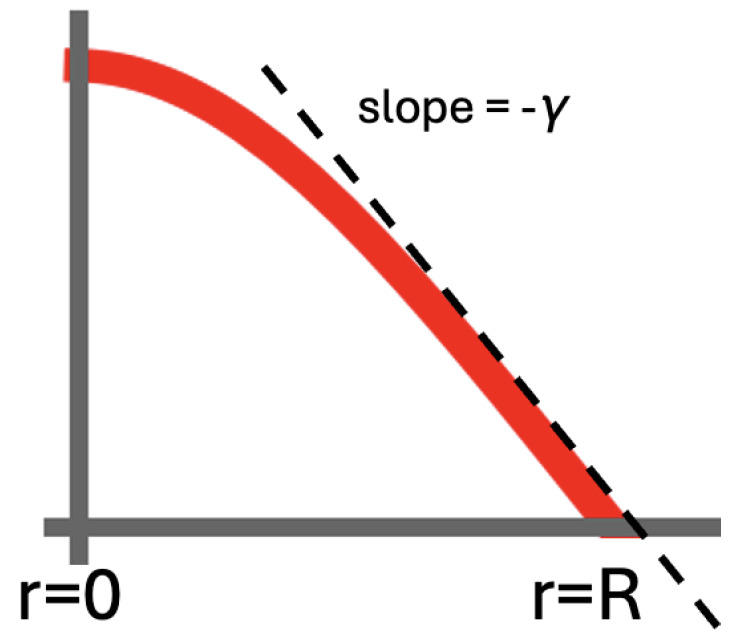
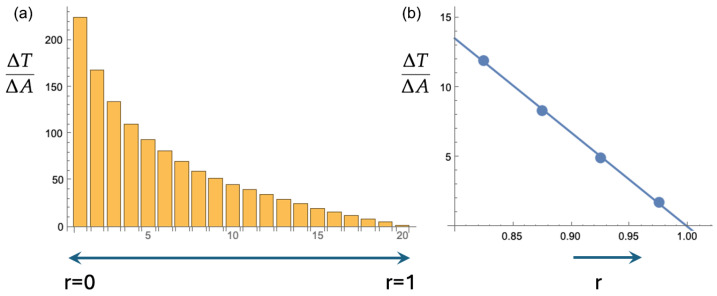
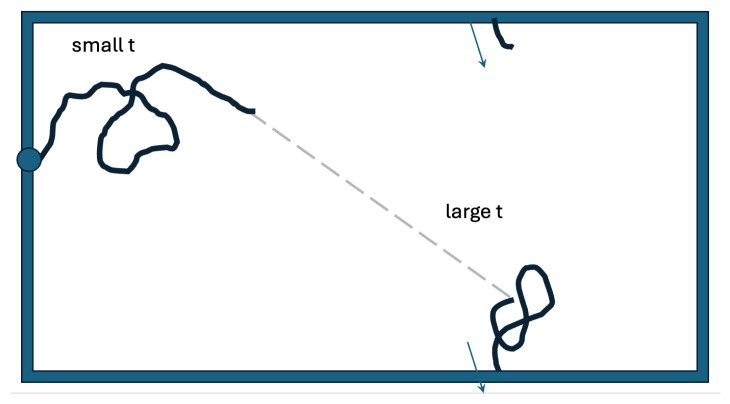
Mammalian fertilization depends on sperm successfully navigating a spatially and chemically complex microenvironment in the female reproductive tract. This process is often conceptualized as a competitive race, but is better understood as a collective random search. Sperm within an ejaculate exhibit a diverse distribution of motility patterns, with some moving in relatively straight lines and others following tightly turning trajectories. Here, we present a two-state random walk model in which sperm switch from high-persistence-length to low-persistence-length motility modes. In reproductive biology, such a switch is often recognized as "hyperactivation". We study a circularly symmetric setup with sperm emerging at the center and searching a finite-area disk. We explore the implications of switching on search efficiency. The first proposed model describes an adaptive search strategy in which sperm achieve improved spatial coverage without cell-to-cell or environment-to-cell communication. The second model that we study adds a small amount of environment-to-cell communication. The models resemble macroscopic search-and-rescue tactics, but without organization or networked communication. Our findings provide a quantitative framework linking sperm motility patterns to efficient search strategies, offering insights into sperm physiology and the stochastic search dynamics of self-propelled particles.
Keywords: persistence length; search trajectories; sperm cell motion.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures

References
-
- Domb C. On multiple returns in the random-walk problem. Math. Proc. Camb. Philos. Soc. 1954;50:586–591. doi: 10.1017/S0305004100029716. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
