

Hydrophobic residue substitutions enhance the stability and in vivo immunogenicity of respiratory syncytial virus fusion protein
- PMID: 40434102
- PMCID: PMC12172430
- DOI: 10.1128/jvi.00087-25
Hydrophobic residue substitutions enhance the stability and in vivo immunogenicity of respiratory syncytial virus fusion protein
Abstract
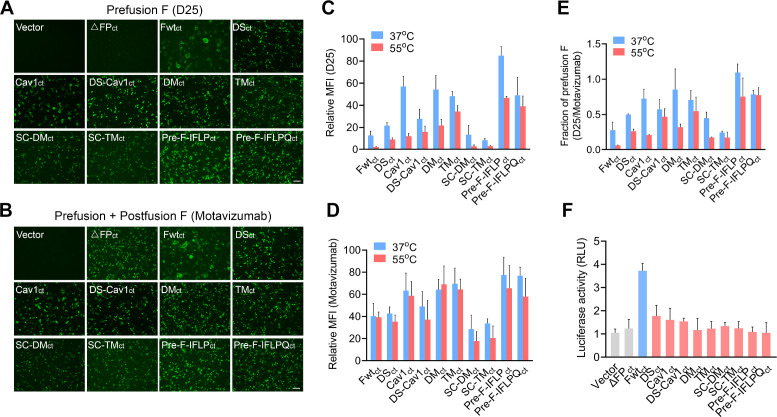
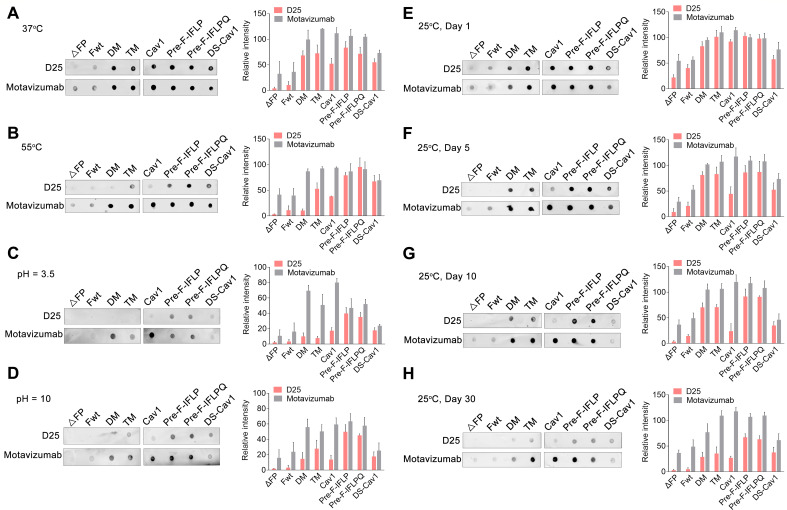
Respiratory syncytial virus (RSV) entry into host cells is facilitated by viral fusion, wherein the metastable RSV fusion (F) protein undergoes a conformational change from a prefusion state to a highly stable postfusion structure. The prefusion F elicits a more robust human antibody response than its postfusion F and is a primary target for RSV vaccine development. However, the inherent instability of the prefusion F trimer and its low protein expression level in host cells are a significant challenge for developing a high-potency RSV vaccine. Here, we report that the introduction of four hydrophobic residue substitutions in the RSV F protein resulted in a highly stable prefusion F trimer (pre-F-IFLP). This engineered variant exhibits enhanced expression and stability compared to DS-Cav1, with improved thermal stability, increased resistance to acid and base, and extended storage life. Furthermore, pre-F-IFLP induced neutralizing antibody responses 72-fold higher than those elicited by DS-Cav1 following a second booster immunization and fully protected mice against RSV infection.
Importance: In this study, we demonstrate that introducing four hydrophobic residue substitutions into the RSV F protein leads to the generation of a highly stable prefusion F trimer (pre-F-IFLP) with improved expression levels in cultured cells and superior stability compared to DS-Cav1, the first-generation prefusion F-stabilized RSV vaccine. Furthermore, pre-F-IFLP induced significantly higher neutralizing antibody responses than DS-Cav1 following both the first and second booster immunizations and conferred complete protection against RSV infection in a mouse model. These findings present an alternative approach for stabilizing the trimeric prefusion F protein, enhancing its expression, and significantly improving its protective efficacy for the prevention of RSV infection in vivo.
Keywords: prefusion F; respiratory syncytial virus; vaccine; viral fusion.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Shi T, McAllister DA, O’Brien KL, Simoes EAF, Madhi SA, Gessner BD, Polack FP, Balsells E, Acacio S, Aguayo C, et al. 2017. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet 390:946–958. doi: 10.1016/S0140-6736(17)30938-8 - DOI - PMC - PubMed
-
- Nair H, Nokes DJ, Gessner BD, Dherani M, Madhi SA, Singleton RJ, O’Brien KL, Roca A, Wright PF, Bruce N, Chandran A, Theodoratou E, Sutanto A, Sedyaningsih ER, Ngama M, Munywoki PK, Kartasasmita C, Simões EAF, Rudan I, Weber MW, Campbell H. 2010. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: a systematic review and meta-analysis. Lancet 375:1545–1555. doi: 10.1016/S0140-6736(10)60206-1 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 82293681(82293680)/National Natural Science Foundation of China
- 82321004/National Natural Science Foundation of China
- 82003610/National Natural Science Foundation of China
- 2022B1515120015, 2024B1515040014/Guangdong Basic and Applied Basic Research Foundation
- 2023B0303000026/Guangdong Major Project of Basic and Applied Basic Research
LinkOut - more resources
Full Text Sources
Medical
