

Protein-primed homopolymer synthesis by an antiviral reverse transcriptase
- PMID: 40436039
- PMCID: PMC12483538
- DOI: 10.1038/s41586-025-09179-5
Protein-primed homopolymer synthesis by an antiviral reverse transcriptase
Abstract
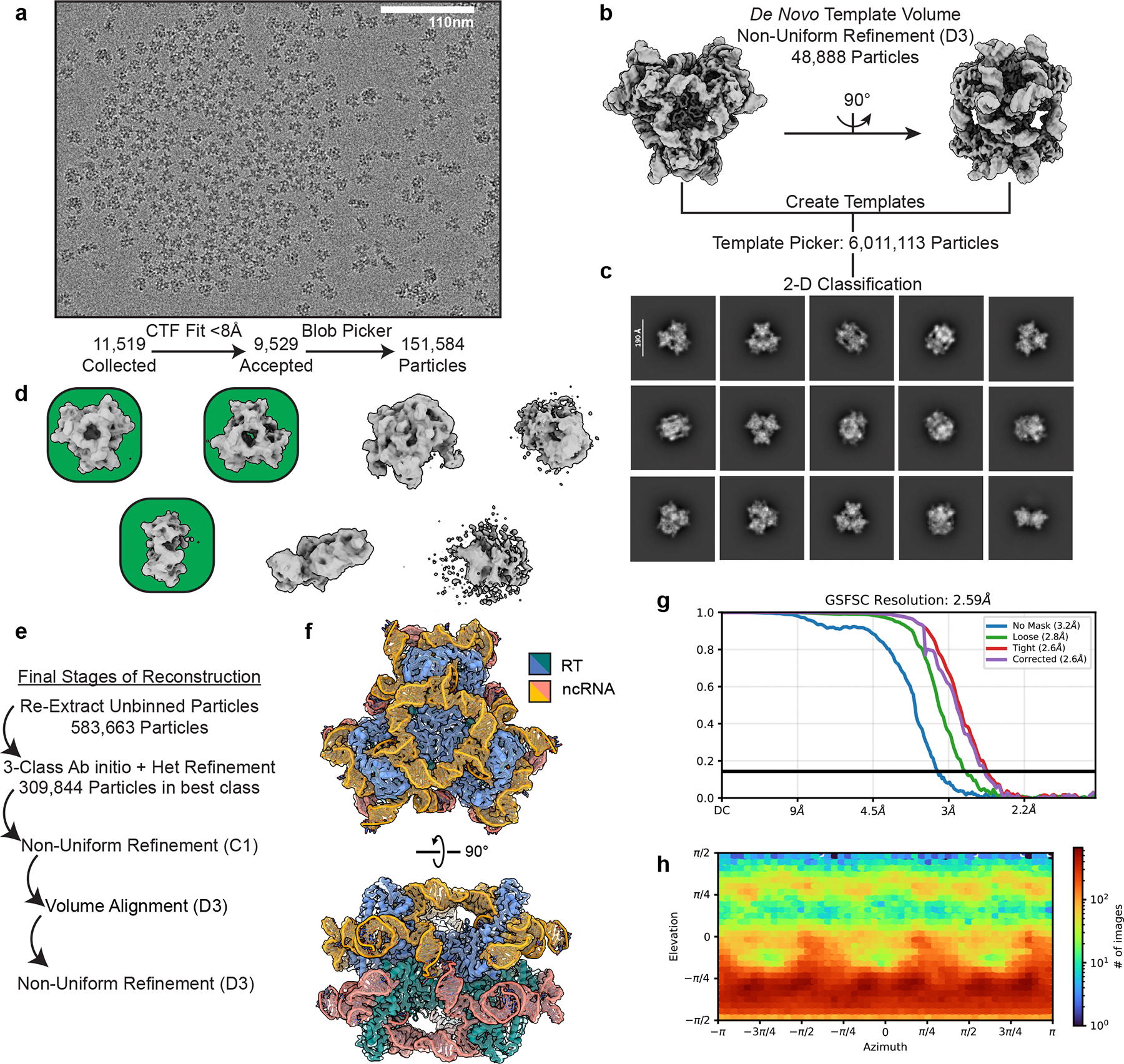
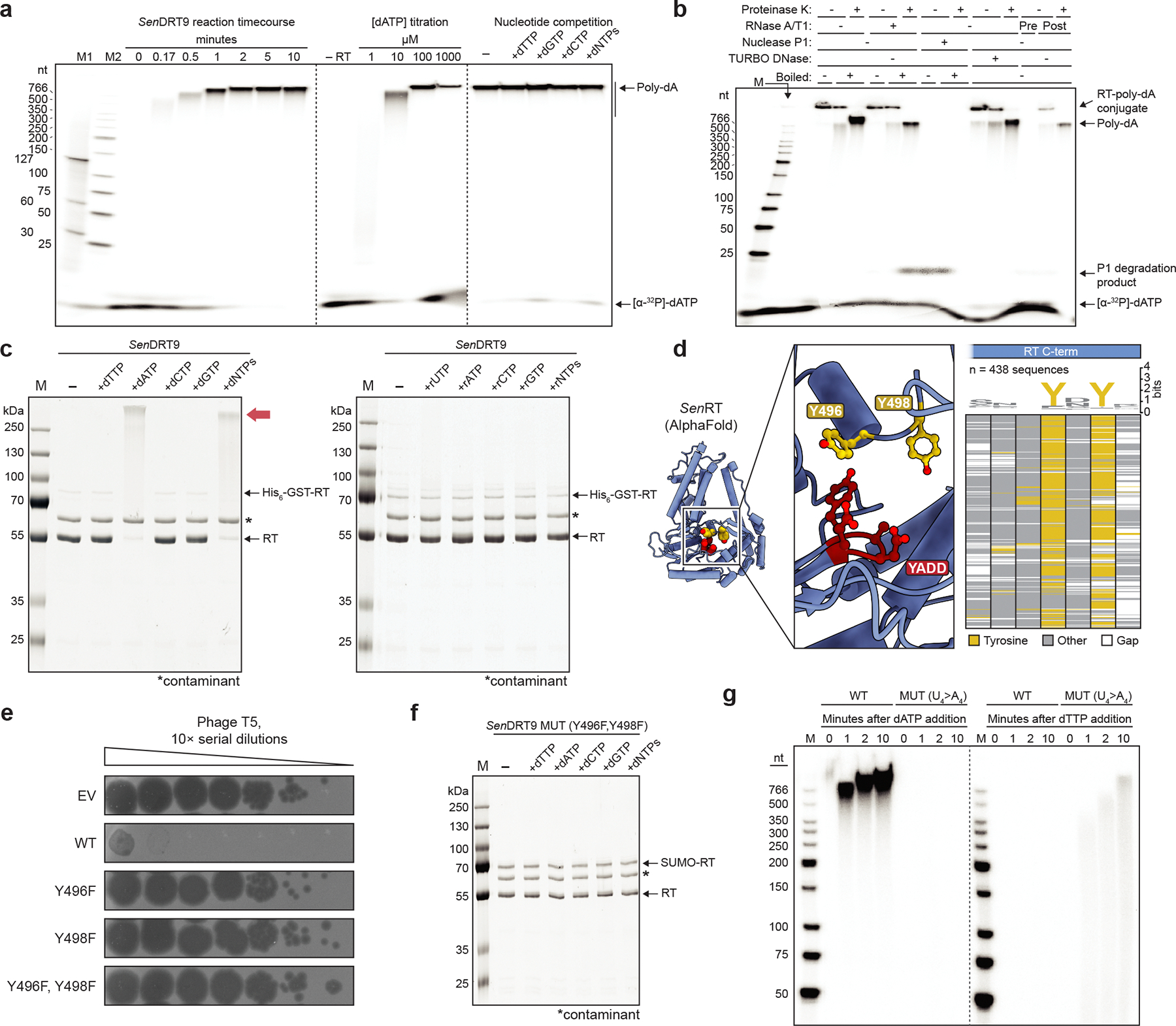
Bacteria defend themselves from viral predation using diverse immune systems, many of which target foreign DNA for degradation1. Defence-associated reverse transcriptase (DRT) systems provide an intriguing counterpoint to this strategy by using DNA synthesis instead2,3. We and others recently showed that DRT2 systems use an RNA template to assemble a de novo gene that encodes the antiviral effector protein Neo4,5. It remains unclear whether similar mechanisms of defence are used by other related DRT families. Here, we show that DRT9 systems defend against phage using DNA homopolymer synthesis. Viral infection triggers polydeoxyadenylate (poly-dA) accumulation in the cell, driving abortive infection and population-level immunity. Cryo-electron microscopy structures reveal how a non-coding RNA serves as both a structural scaffold and reverse transcription template to direct hexameric complex assembly and poly-dA synthesis. Notably, biochemical and functional experiments identify tyrosine residues within the reverse transcriptase itself that probably prime DNA synthesis, leading to the formation of protein-DNA covalent adducts. Synthesis of poly-dA by DRT9 in vivo is regulated by the competing activities of phage-encoded triggers and host-encoded silencers. Collectively, our study identifies a nucleic-acid-driven defence system that expands the paradigm of bacterial immunity and broadens the known functions of reverse transcriptases.
© 2025. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests: S.H.S. is a co-founder and scientific advisor to Dahlia Biosciences, a scientific advisor to CrisprBits and Prime Medicine, and an equity holder in Dahlia Biosciences and CrisprBits. B.W. is the founder of SurGene and is listed as an inventor on patent applications related to CRISPR–Cas systems and applications thereof. The other authors declare no competing interests.
Figures













Update of
-
Protein-primed DNA homopolymer synthesis by an antiviral reverse transcriptase.bioRxiv [Preprint]. 2025 Mar 25:2025.03.24.645077. doi: 10.1101/2025.03.24.645077. bioRxiv. 2025. Update in: Nature. 2025 Jul;643(8074):1352-1362. doi: 10.1038/s41586-025-09179-5. PMID: 40196691 Free PMC article. Updated. Preprint.
References
MAIN TEXT REFERENCES
-
- Georjon H & Bernheim A The highly diverse antiphage defence systems of bacteria. Nat. Rev. Microbiol. 21, 686–700 (2023). - PubMed
ADDITIONAL REFERENCES
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
