

WNK1 mediates M-CSF-induced macropinocytosis to enforce macrophage lineage fidelity
- PMID: 40436823
- PMCID: PMC12120055
- DOI: 10.1038/s41467-025-59901-0
WNK1 mediates M-CSF-induced macropinocytosis to enforce macrophage lineage fidelity
Abstract
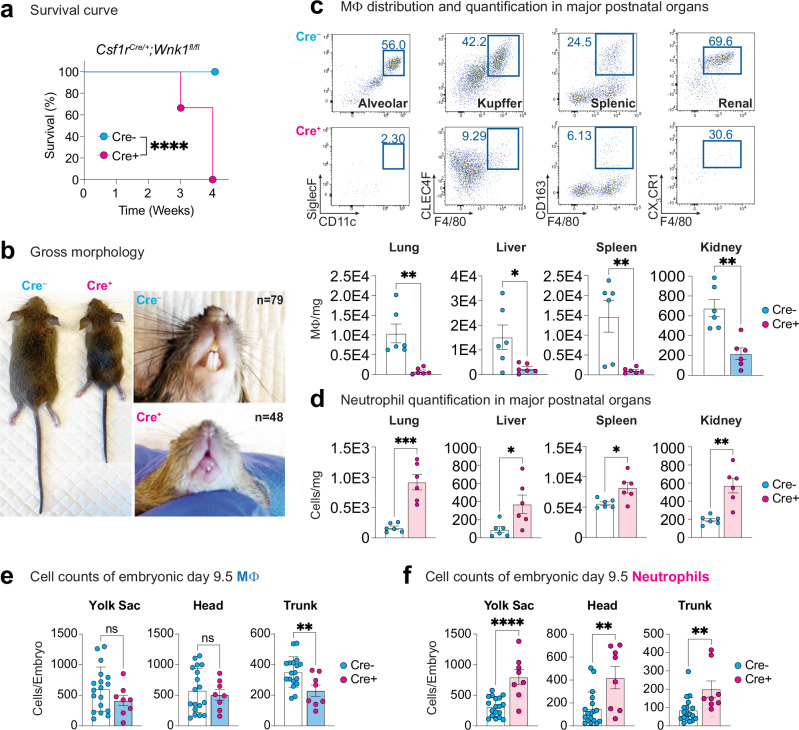
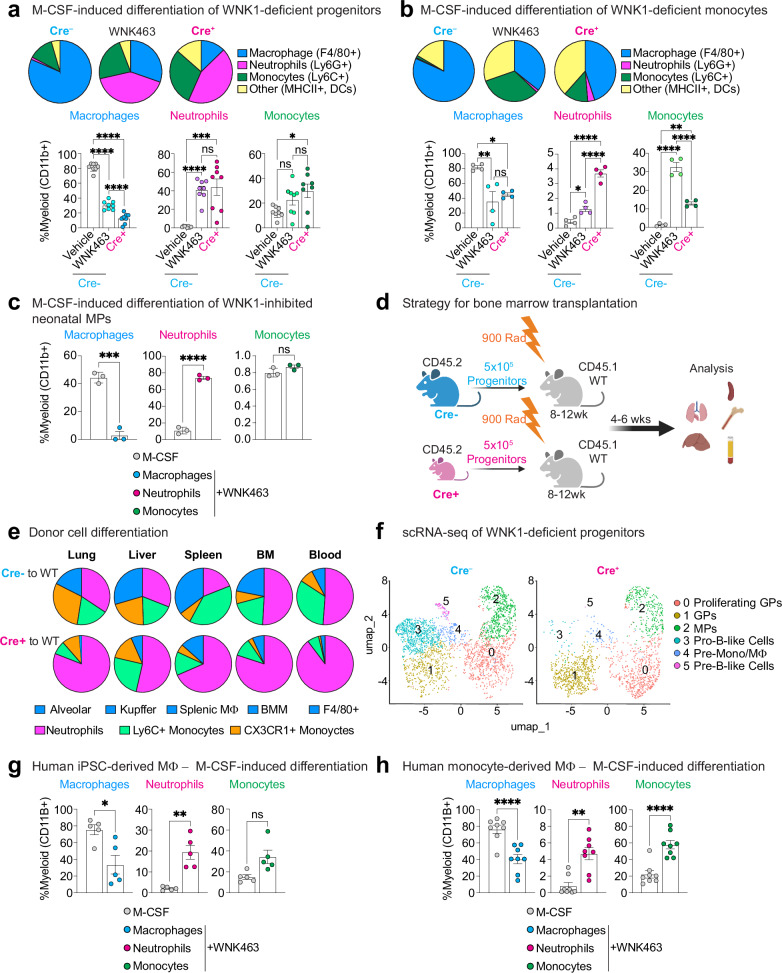
Tissue-resident macrophages (TRM) are critical for mammalian organismal development and homeostasis. Here we report that with-no-lysine 1 (WNK1) controls myeloid progenitor fate, with Csf1riCre-mediated Wnk1 deletion in mice (WNK1-deficient mice) resulting in loss of TRMs and causing perinatal mortality. Mechanistically, absence of WNK1 or inhibition of WNK kinase activity disrupts macrophage colony-stimulating factor (M-CSF)-stimulated macropinocytosis, thereby blocking mouse and human progenitor and monocyte differentiation into macrophages and skewing progenitor differentiation into neutrophils. Treatment with PMA rescues macropinocytosis but not macrophage differentiation of WNK-inhibited progenitors, implicating that M-CSF-stimulated, macropinocytosis-induced activation of WNK1 is required for macrophage differentiation. Finally, M-CSF-stimulated macropinocytosis triggers WNK1 nuclear translocation and concomitant increased protein expression of interferon regulatory factor (IRF)8, whereas inhibition of macropinocytosis or WNK kinase activity suppresses IRF8 expression. Our results thus suggest that WNK1 and downstream IRF8-regulated genes are important for M-CSF/macropinocytosis-mediated regulation of myeloid cell lineage commitment during TRM development and homeostasis.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: J.S.A.P. holds patents related to imaging and modulation of cellular metabolism stemming from this work and is a co-founder of Atish Technologies. All other authors have no competing interests to disclose.
Figures



References
MeSH terms
Substances
Grants and funding
- R00 CA237728/CA/NCI NIH HHS/United States
- DP2 GM146337/GM/NIGMS NIH HHS/United States
- 5T32CA009149/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- K08 MH130773/MH/NIMH NIH HHS/United States
- MR/W019264/1/RCUK | Medical Research Council (MRC)
- U19 AI135990/AI/NIAID NIH HHS/United States
- T32 CA009149/CA/NCI NIH HHS/United States
- K99 CA237728/CA/NCI NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- R01DK098145/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- K99 HL171844/HL/NHLBI NIH HHS/United States
- 5U54CA209975/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- DK111542/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- 1K99HL171844/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- Research Scholars Award/Pew Charitable Trusts
- L30 CA274750/CA/NCI NIH HHS/United States
- 5R00CA237728/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- MR/X019314/1/RCUK | Medical Research Council (MRC)
- U54 DK137329/DK/NIDDK NIH HHS/United States
- P30CA008748/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- 5K08MH130773/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 DK111542/DK/NIDDK NIH HHS/United States
- U54 CA209975/CA/NCI NIH HHS/United States
- U54DK137329/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 DK098145/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
