

Two cancer cell lines utilize Myosin 10 and the kinesin HSET differentially to maintain mitotic spindle bipolarity
- PMID: 40440343
- PMCID: PMC12121739
- DOI: 10.1371/journal.pone.0325016
Two cancer cell lines utilize Myosin 10 and the kinesin HSET differentially to maintain mitotic spindle bipolarity
Abstract
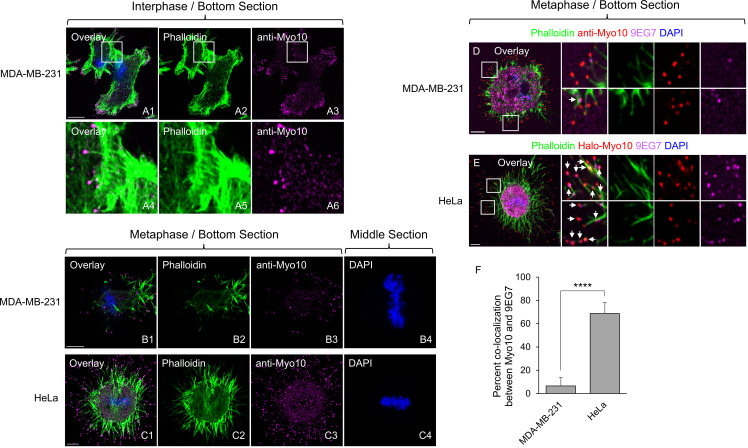
Cancer cells often undergo mitosis possessing more than two centrosomes. To avoid a multipolar mitosis, the consequences of which are typically aneuploidy induced senescence, they must cluster their extra centrosomes to create a pseudo-bipolar spindle. Such supernumerary centrosome clustering (SNCC) requires Myosin 10 (Myo10) and the pole-focusing kinesin HSET. We showed recently that Myo10 promotes SNCC in HeLa cells by promoting retraction fiber-based cell adhesion, and that it further supports spindle bipolarity by preventing the generation of extra spindle poles via pericentriolar material (PCM) fragmentation. Here we quantified the contribution that Myo10 and HSET make individually and together to SNCC and PCM/pole integrity in HeLa cells and in MDA-MB-231 cells, which differ from HeLa in being more dependent on SNCC and less dependent on retraction fiber-based cell adhesion. As expected, knockdown of Myo10 and HSET individually increased the frequency of multipolar spindles in both cell types. Their effects were surprisingly not additive, however. For HeLa and MDA-MB-231 cells undergoing mitosis with more than two centrosomes, the defect in SNCC was almost entirely responsible for their multipolar phenotype following knockdown of either Myo10 or HSET. For HeLa and MDA-MB-231 cells undergoing mitosis with two centrosomes, PCM/pole fragmentation was the primary cause of multipolar spindles following HSET knockdown. Unlike HeLa, however, MDA-MB-231 cells exhibited very little PCM/pole fragmentation following Myo10 knockdown. This difference may be due to the smaller role that Myo10 plays in retraction fiber-based adhesion in MDA-MB-231. Finally, we show that HSET knockdown disrupts retraction fiber formation and organization, which may explain why the defects in double knockdown cells were not significantly greater than in HSET knockdown cells. These and other results can inform efforts to target these two motor proteins to selectively kill cancer cells by increasing their frequency of multipolar divisions.
Copyright: This is an open access article, free of all copyright, and may be freely reproduced, distributed, transmitted, modified, built upon, or otherwise used by anyone for any lawful purpose. The work is made available under the Creative Commons CC0 public domain dedication.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
