

Distinct viral reservoirs and immune signatures in individuals on long-term antiretroviral therapy with perinatally acquired HIV-1
- PMID: 40446801
- PMCID: PMC12208329
- DOI: 10.1016/j.xcrm.2025.102150
Distinct viral reservoirs and immune signatures in individuals on long-term antiretroviral therapy with perinatally acquired HIV-1
Abstract
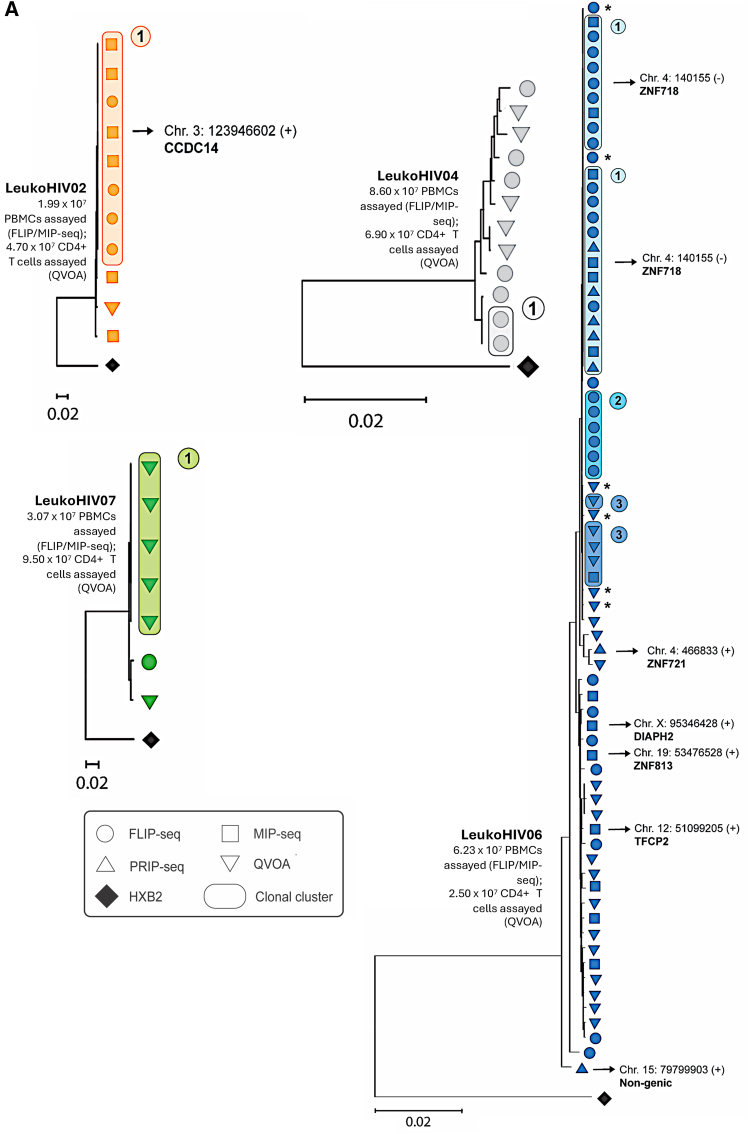
Early initiation of antiretroviral therapy (ART) following HIV-1 infection restricts the size of the latent reservoir, following both horizontal and vertical infections. Here, we comprehensively profile the reservoirs and immunological milieus of nine young adults who acquired HIV-1 perinatally and remained on suppressive long-term ART (median: 20 years) since infancy (LeukoHIV cohort). Genome-intact reservoirs are markedly smaller compared to a cohort of adults on suppressive ART started in adulthood, with some LeukoHIV individuals characterized by an absence or near absence of intact proviruses in up to a billion peripheral blood mononuclear cells (PBMCs). Higher frequencies of functional CD56bright natural killer (NK) cells with increased cytotoxic activity are detectable in the LeukoHIV cohort compared to an adult reference cohort, while one LeukoHIV participant displayed a potent HIV-1-specific CD8+ T cell response. Collectively, our data suggest that long-term ART initiated in early life following perinatal transmission may facilitate an immune environment better equipped to restrict the HIV-1 reservoir.
Keywords: HIV; MIP-seq; children; deep latency; leukophoresis; long-term ART; natural killer cells; perinatal infection; provirus; reservoir.
Copyright © 2025 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests N. Cotugno, P.P., and G.R.P. are respectively CEO and co-founders at Probiomics SRL.
Figures





References
-
- HIV/AIDS GJUNPo. UNAIDS Data 2023. 2023.
-
- Chi B.H., Adler M.R., Bolu O., Mbori-Ngacha D., Ekouevi D.K., Gieselman A., Chipato T., Luo C., Phelps B.R., McClure C., et al. Progress, Challenges, and New Opportunities for the Prevention of Mother-to-Child Transmission of HIV Under the US President's Emergency Plan for AIDS Relief. J. Acquir. Immune Defic. Syndr. 2012;60:S78–S87. - PMC - PubMed
-
- UNAIDS/WHO. Epidermiological fact sheets on HIV/AIDS and Sexually Transmitted Infections, 2004 Update. 2004.
-
- Boerma R.S., Boender T.S., Bussink A.P., Calis J.C.J., Bertagnolio S., Rinke de Wit T.F., Boele van Hensbroek M., Sigaloff K.C.E. Suboptimal Viral Suppression Rates Among HIV-Infected Children in Low- and Middle-Income Countries: A Meta-analysis. Clin. Infect. Dis. 2016;63:1645–1654. - PubMed
-
- Han W.M., Law M.G., Egger M., Wools-Kaloustian K., Moore R., McGowan C., Kumarasamy N., Desmonde S., Edmonds A., Davies M.A., et al. Global estimates of viral suppression in children and adolescents and adults on antiretroviral therapy adjusted for missing viral load measurements: a multiregional, retrospective cohort study in 31 countries. Lancet. HIV. 2021;8:e766–e775. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
