

Post-fertilization transcription initiation in an ancestral LTR retrotransposon drives lineage-specific genomic imprinting of ZDBF2
- PMID: 40455561
- PMCID: PMC12129452
- DOI: 10.7554/eLife.94502
Post-fertilization transcription initiation in an ancestral LTR retrotransposon drives lineage-specific genomic imprinting of ZDBF2
Abstract
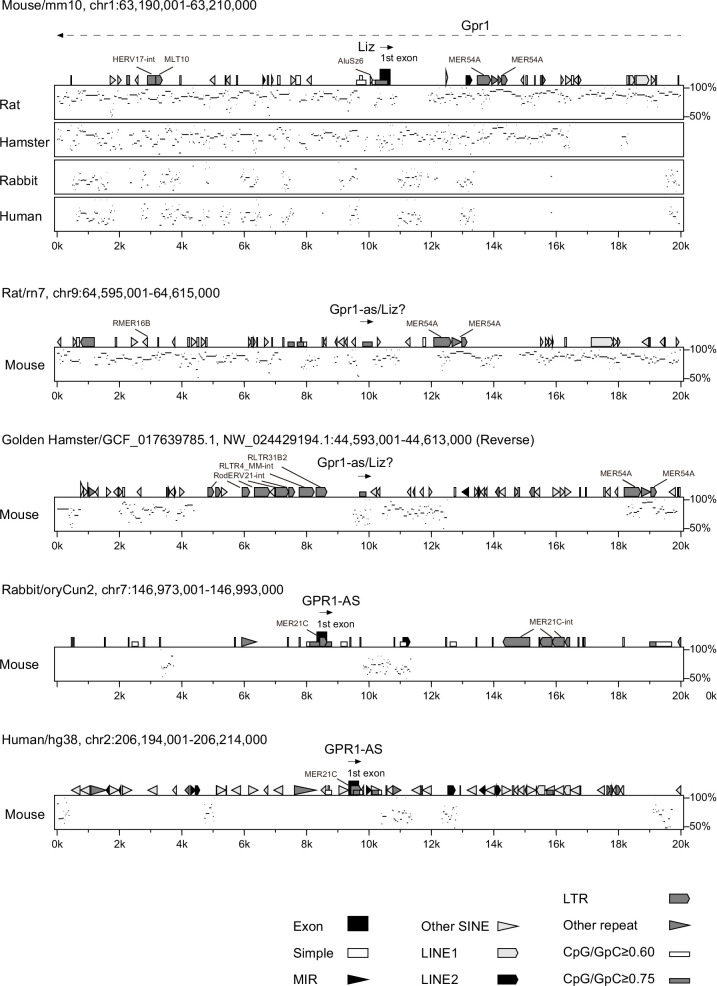
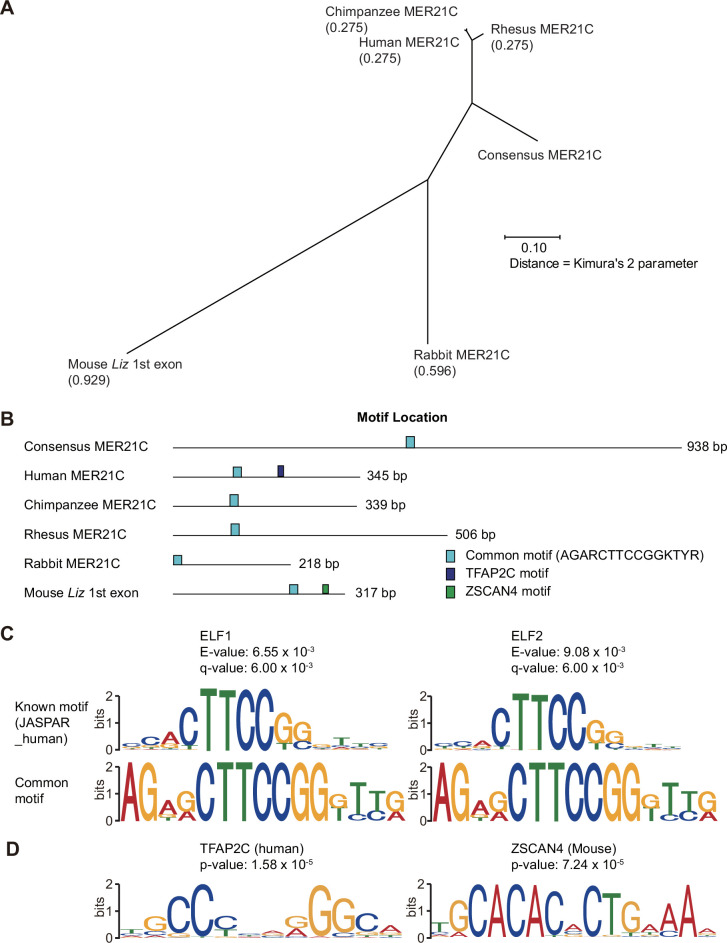
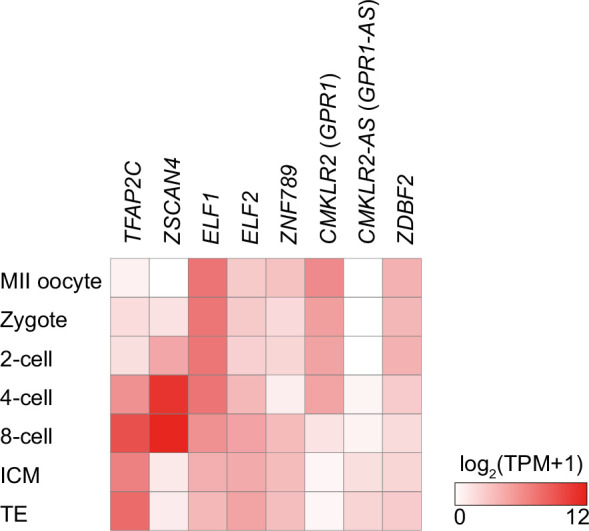
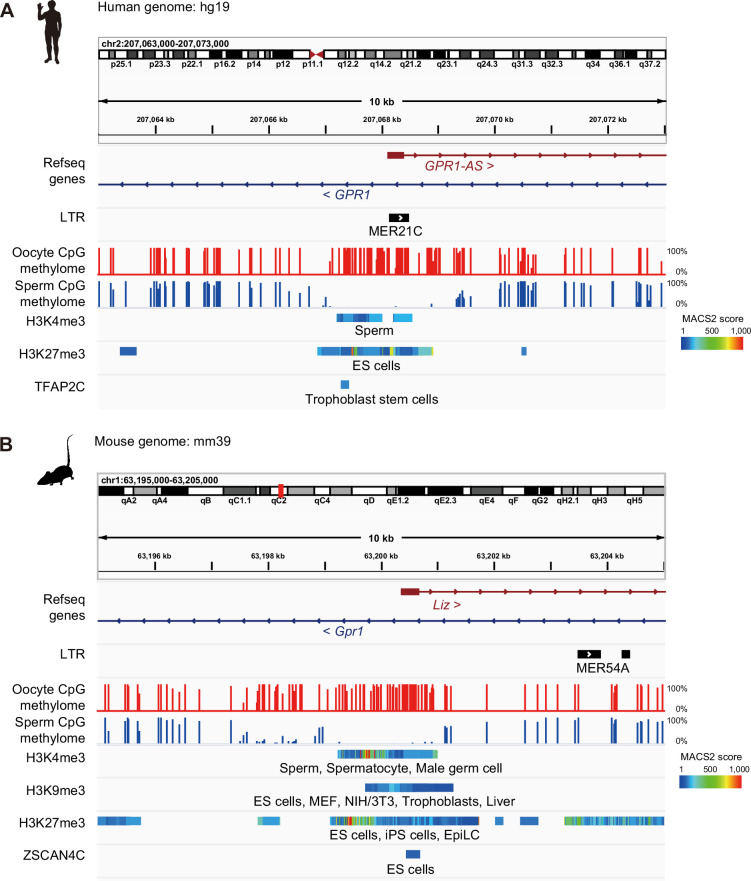
The imprinted gene ZDBF2 is regulated through a unique mechanism involving a transient paternal transcript in early embryos, rather than persistent gametic DNA methylation. In humans and mice, this transcript-CMKLR2-AS (also known as GPR1-AS) or the long isoform of Zdbf2 (Liz/Zdbf2linc/Platr12)-arises from the unmethylated paternal allele and initiates secondary epigenetic marks that maintain ZDBF2 expression. Here, we investigate the evolutionary origin of this mechanism, and show that the first exon of human GPR1-AS overlaps with a MER21C long terminal repeat (LTR), a retrotransposon subfamily specific to Boreoeutherian mammals. Comparative analyses revealed that this MER21C insertion occurred in the common ancestor of Euarchontoglires, including primates, rodents, and rabbits. Although not annotated, the first exon of mouse Liz displays conserved features with the MER21C-overlapping exon in humans. In rabbit and nonhuman primate placentas, GPR1-AS orthologs with LTR-embedded first exons were also identified. In contrast, in non-Euarchontoglire mammals such as cow and tammar wallaby, ZDBF2 is biallelically expressed, suggesting absence of imprinting. These findings suggest that ZDBF2 imprinting emerged in Euarchontoglires via MER21C insertion. Together with our prior work on LTR-driven imprinting in oocytes, our findings demonstrate that post-fertilization activation of retrotransposons can also drive lineage-specific acquisition of imprinting.
Keywords: LTR retrotransposon; RNA-seq; chromosomes; evolutionary biology; gene expression; genomic imprinting; human; mammalian evolution; mouse; placenta; rhesus macaque.
© 2024, Kobayashi et al.
Conflict of interest statement
HK, TI, SK, KT, TT, SN, SS, MH, MR, MK, SS, TK, HN, HM, KN, AU, HK, MK, HI, KN, ML, KK No competing interests declared
Figures













Update of
- doi: 10.1101/2023.10.30.564869
- doi: 10.7554/eLife.94502.1
- doi: 10.7554/eLife.94502.2
References
-
- Armstrong DL, McGowen MR, Weckle A, Pantham P, Caravas J, Agnew D, Benirschke K, Savage-Rumbaugh S, Nevo E, Kim CJ, Wagner GP, Romero R, Wildman DE. The core transcriptome of mammalian placentas and the divergence of expression with placental shape. Placenta. 2017;57:71–78. doi: 10.1016/j.placenta.2017.04.015. - DOI - PMC - PubMed
-
- Armstrong J, Hickey G, Diekhans M, Fiddes IT, Novak AM, Deran A, Fang Q, Xie D, Feng S, Stiller J, Genereux D, Johnson J, Marinescu VD, Alföldi J, Harris RS, Lindblad-Toh K, Haussler D, Karlsson E, Jarvis ED, Zhang G, Paten B. Progressive Cactus is a multiple-genome aligner for the thousand-genome era. Nature. 2020;587:246–251. doi: 10.1038/s41586-020-2871-y. - DOI - PMC - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- SRA/ERR489144
- SRA/SRR543893
- SRA/SRR558617
Grants and funding
- Grant-in-Aid for Scientific Research (B) JP21H02382/Japan Society for the Promotion of Science
- Grant-in-Aid for Scientific Research (B) JP23K21282/Japan Society for the Promotion of Science
- Grant-in-Aid for Scientific Research (B) JP25K02190/Japan Society for the Promotion of Science
- Grant-in-Aid for Scientific Research (A) JP20H00471/Japan Society for the Promotion of Science
- Grant No. 202310022/Mitsubishi Foundation
LinkOut - more resources
Full Text Sources
