

Effect of variable phosphorus availability on root mechanisms involved in mobilization of the soil P in three lupine species
- PMID: 40456812
- PMCID: PMC12130325
- DOI: 10.1038/s41598-025-03694-1
Effect of variable phosphorus availability on root mechanisms involved in mobilization of the soil P in three lupine species
Abstract
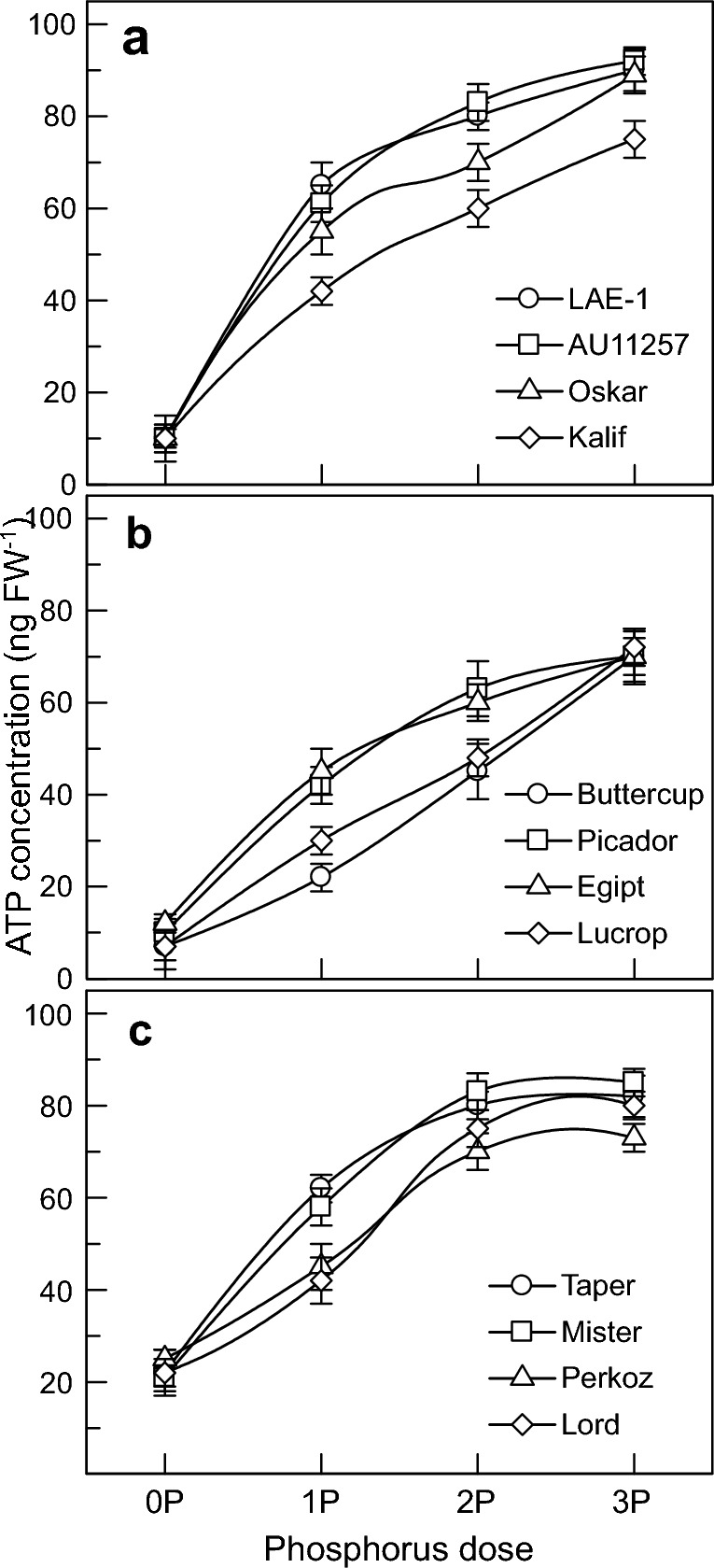
Improving the soil phosphorus (P) acquisition efficiency by plants is one of the most important challenges for modern agriculture. Only 15-20% of this nutrient applied to the soil with fertilisers is used by plants, with the majority being converted to less available forms. Lupine species are a great genetic model system with significant potential to provide novel information. Here we present an investigation into the effects of variable availability in the rhizosphere on P-mobilising mechanisms and photosynthetic activity, studied in 12 varieties of three lupine species. P uptake was significantly stimulated by low molecular weight organic acid (OA) exudation and H+-ATPase-mediated proton transport in roots. The predominant mechanism in white lupine was the H+ release into the rhizosphere, OA exudation predominated in yellow lupine, while narrow-leaved lupine varieties used both strategies effectively. Three lupine species showed significant differences in the quantitative and qualitative composition of carboxylic acids in root exudates. The variable P availability in soil solution significantly affected the photosynthetic performance of the plants studied. At the same time, the activity of the photosynthetic electron transport chain and photosynthetic CO2 assimilation was a key factor determining activity of the mechanisms involved in P mobilisation. We discuss the implications of these contrasting strategies for our understanding of tolerance to low P and in relation to breeding plants with higher P acquisition efficiency.
Keywords: H+-ATPase; Low-molecular-weight organic acids; Lupines; Phosphorus availability; Photosynthetic activity.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interests: The authors declare no competing interests.
Figures






 citric acid,
citric acid,  malic acid,
malic acid,  acetic acid and
acetic acid and  succinic acid. The plants were supplemented with 0 (0P; control), 22 (1P), 44 (2P) and 88 (3P) mg kg−1 of P. Data points represent the means ± SE (n = 4). The same letter denote homogenous groups (interaction P dose × variety) within species after two-way ANOVA and Bonferroni post hoc test at p = 0.05.
succinic acid. The plants were supplemented with 0 (0P; control), 22 (1P), 44 (2P) and 88 (3P) mg kg−1 of P. Data points represent the means ± SE (n = 4). The same letter denote homogenous groups (interaction P dose × variety) within species after two-way ANOVA and Bonferroni post hoc test at p = 0.05.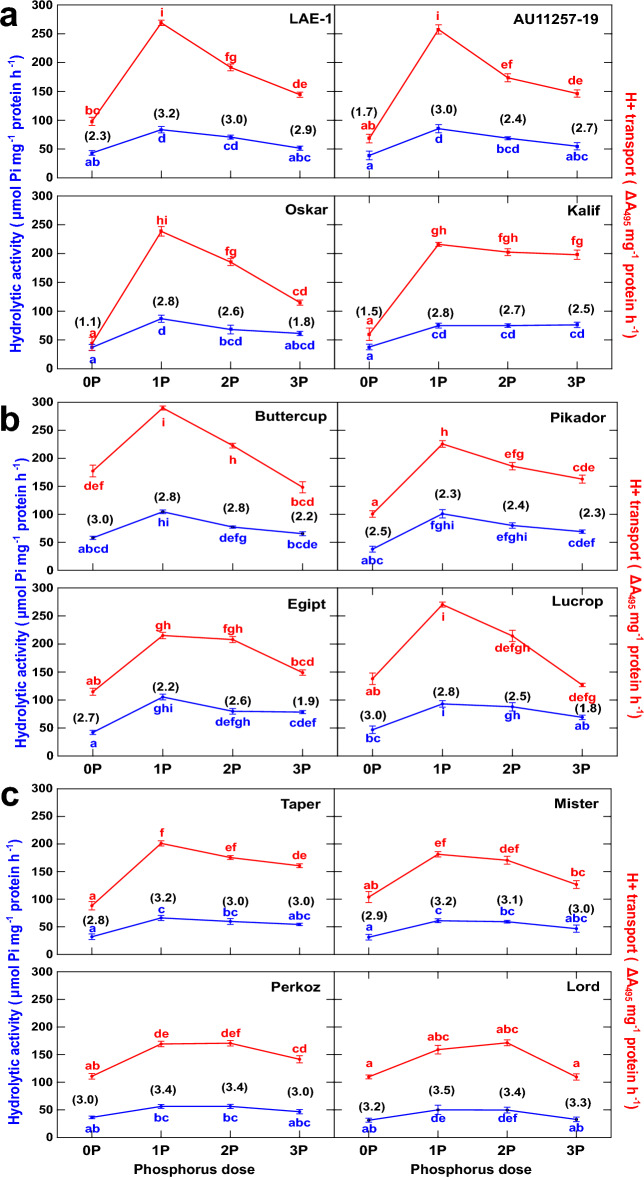
 ) and the H+-ATPase-mediated proton efflux (
) and the H+-ATPase-mediated proton efflux ( ) in roots of narrow-leafed (a), white (b) and yellow (c) lupines, grown under variable P supplementation. The plants were supplemented with 0 (0P; control), 22 (1P), 44 (2P) and 88 (3P) mg kg−1 of P. The values of the H+/ATP coupling ratio are presented in brackets. Data points represent the means ± SE (n = 4). The same letter denote homogenous groups (interaction P dose × variety) within species after two-way ANOVA and Bonferroni post hoc test at p = 0.05.
) in roots of narrow-leafed (a), white (b) and yellow (c) lupines, grown under variable P supplementation. The plants were supplemented with 0 (0P; control), 22 (1P), 44 (2P) and 88 (3P) mg kg−1 of P. The values of the H+/ATP coupling ratio are presented in brackets. Data points represent the means ± SE (n = 4). The same letter denote homogenous groups (interaction P dose × variety) within species after two-way ANOVA and Bonferroni post hoc test at p = 0.05.


Similar articles
-
Root exudation, phosphorus acquisition, and microbial diversity in the rhizosphere of white lupine as affected by phosphorus supply and atmospheric carbon dioxide concentration.J Environ Qual. 2005 Nov 7;34(6):2157-66. doi: 10.2134/jeq2004.0423. Print 2005 Nov-Dec. J Environ Qual. 2005. PMID: 16275716
-
Influence of root exudation of white lupine (Lupinus albus L.) on uranium phytoavailability in a naturally uranium-rich soil.J Environ Radioact. 2018 Oct;190-191:39-50. doi: 10.1016/j.jenvrad.2018.04.022. Epub 2018 May 15. J Environ Radioact. 2018. PMID: 29751206
-
Time and substrate dependent exudation of carboxylates by Lupinus albus L. and Brassica napus L.Plant Physiol Biochem. 2011 Nov;49(11):1272-8. doi: 10.1016/j.plaphy.2011.08.012. Epub 2011 Sep 14. Plant Physiol Biochem. 2011. PMID: 22000050
-
Enhancing phosphorus and zinc acquisition efficiency in rice: a critical review of root traits and their potential utility in rice breeding.Ann Bot. 2013 Jul;112(2):331-45. doi: 10.1093/aob/mcs217. Epub 2012 Oct 15. Ann Bot. 2013. PMID: 23071218 Free PMC article. Review.
-
Physiological and molecular alterations in plants exposed to high [CO2] under phosphorus stress.Biotechnol Adv. 2015 May-Aug;33(3-4):303-16. doi: 10.1016/j.biotechadv.2015.03.011. Epub 2015 Mar 20. Biotechnol Adv. 2015. PMID: 25797341 Review.
References
-
- Marschner, H. Mineral Nutrition of Higher Plants 2nd edn, 889 (Academic Press, 1995).
-
- Hopkins, B. G. Phosphorus. In Handbook of Plant Nutrition 2nd edn (eds Allen, V. & Barker Pilbeam, D. J.) 65–126 (CRC Press Taylor & Francis Group, 2015).
-
- Vance, C. P., Uhde-Stone, C. & Allan, D. L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resources. New Phytol.157, 423–447 (2003). - PubMed
-
- Amadou, I., Houben, D., Lambers, H. & Faucon, M.-P. Key role of root trait combinations and plasticity in response to phosphorus forms on phosphorus-acquisition in agroecosystems. Plant Soil509, 123 (2024).
-
- Amadou, I., Houben, D. & Faucon, M.-P. Unravelling the role of rhizosphere microbiome and root traits in organic phosphorus mobilization for sustainable phosphorus fertilization. A review. Agronomy11(11), 2267 (2021).
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
