

Activation of Human FPR2 with AT-RvD1 Resolves Acute Sialadenitis in Vivo
- PMID: 40464830
- PMCID: PMC12353947
- DOI: 10.1007/s10753-025-02320-6
Activation of Human FPR2 with AT-RvD1 Resolves Acute Sialadenitis in Vivo
Abstract
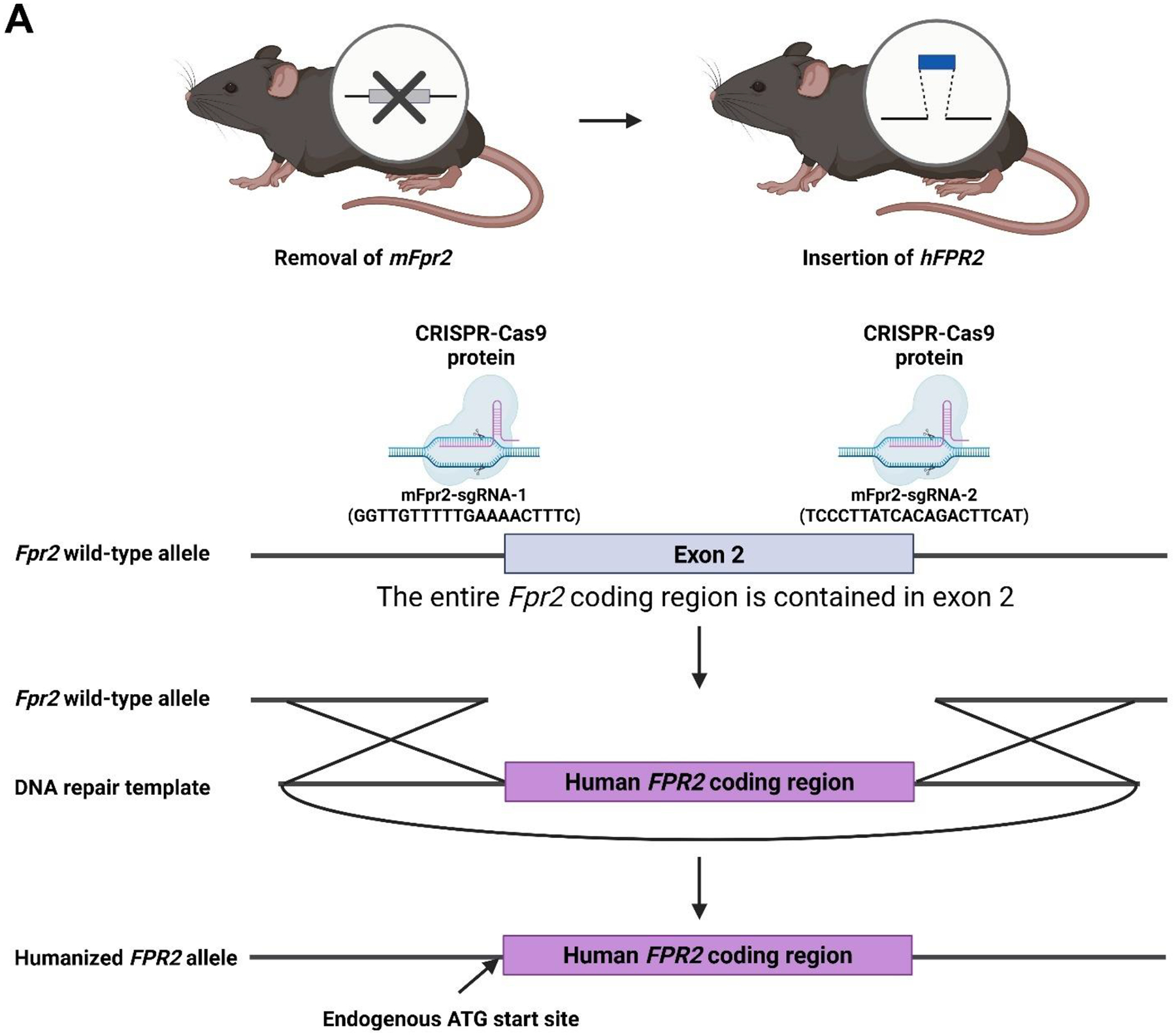
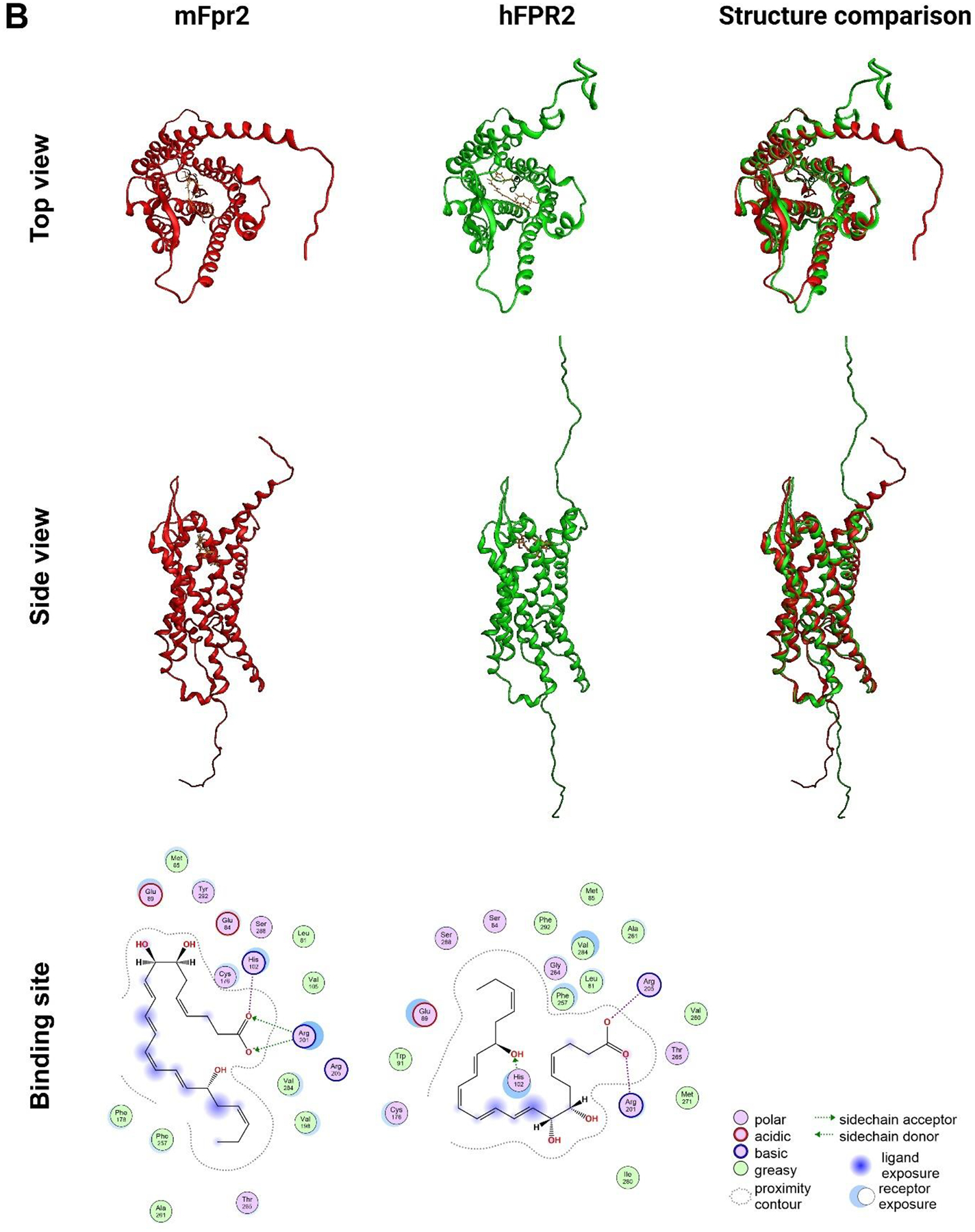
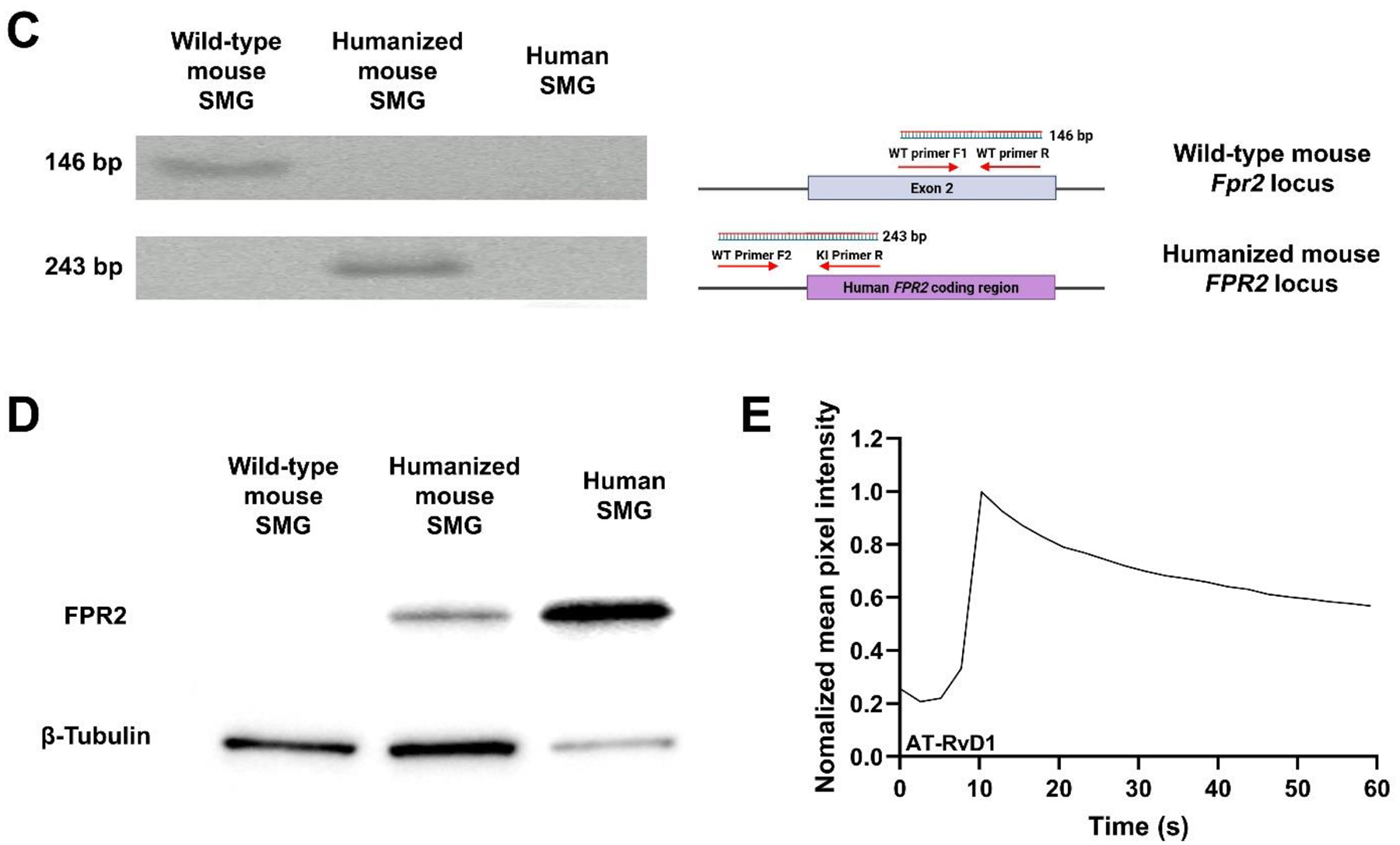
Previous studies demonstrated that activation of the mouse G protein-coupled formyl peptide receptor 2 (mFpr2) with aspirin-triggered resolvin D1 (AT-RvD1) blocks pro-inflammatory cytokine signaling while promoting salivary gland (SG) epithelial integrity both in vitro and in vivo. In addition, mice lacking Fpr2 display alterations of SG innate and adaptive immunity. Taken together, these results indicate that Fpr2 activation with AT-RvD1 restores saliva secretion and regulates SG immunity in mice. To demonstrate the value of AT-RvD1 for use in human SG, however, we need to extend the findings above in the direction of clinical use. To this end, the current study investigated whether treatment with AT-RvD1 reduces SG inflammation and restores saliva secretion in an acute sialadenitis mouse model expressing the human formyl peptide receptor 2 (hFPR2) protein. Results indicate that mice carrying the hFPR2 and treated with lipopolysaccharide (LPS) display acute sialadenitis-like features as shown by increased levels of proliferating inflammatory cells, loss of epithelial integrity and reduced saliva secretion. In contrast, when these mice are treated with AT-RvD1, the sialadenitis-like features are drastically reduced as evidenced by a significant decrease in proliferating inflammatory cells as well as restoration of saliva secretion to levels comparable to phosphate buffered saline (PBS)-treated healthy controls. Finally, changes observed in mice carrying the hFPR2 and treated with LPS and AT-RvD1 were comparable to those observed in wild-type mice carrying the mFpr2. Together, these results demonstrate that activation of hFPR2 with AT-RvD1 resolves acute sialadenitis in vivo.
Keywords: Immunity; Inflammation; Lipids; Oral pathology; Receptors; Salivary physiology.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Competing interest: The authors declare no competing interests. Ethical Approval: All animal handling, anesthesia, and treatments were conducted in compliance with Animal Research: Reporting of In Vivo Experiments guidelines and received prior approval from the University of Missouri-Columbia Animal Care and Use Committee (ACUC), approval number: 42063. Moreover, all human specimen analyses were conducted under the guidelines and with the approval of the University of Missouri-Columbia Institutional Review Board (IRB), approval number: 2024461, with informed consent obtained for each patient collected from their medical record.
Figures




References
-
- McQuone SJ, Acute viral and bacterial infections of the salivary glands. Otolaryngol Clin North Am, 1999. 32(5): p. 793–811. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
