

Single cell profiling of human airway identifies tuft-ionocyte progenitor cells displaying cytokine-dependent differentiation bias in vitro
- PMID: 40467553
- PMCID: PMC12137820
- DOI: 10.1038/s41467-025-60441-w
Single cell profiling of human airway identifies tuft-ionocyte progenitor cells displaying cytokine-dependent differentiation bias in vitro
Abstract
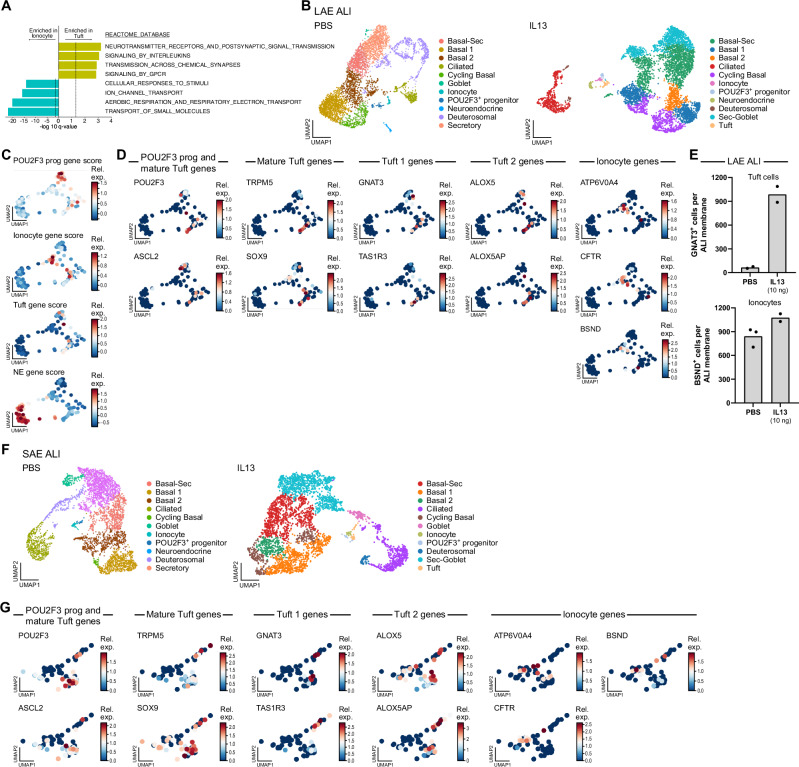
Human airways contain specialized rare epithelial cells including CFTR-rich ionocytes that regulate airway surface physiology and chemosensory tuft cells that produce asthma-associated inflammatory mediators. Here, using a lung cell atlas of 311,748 single cell RNA-Seq profiles, we identify 687 ionocytes (0.45%). In contrast to prior reports claiming a lack of ionocytes in the small airways, we demonstrate that ionocytes are present in small and large airways in similar proportions. Surprisingly, we find only 3 mature tuft cells (0.002%), and demonstrate that previously annotated tuft-like cells are instead highly replicative progenitor cells. These tuft-ionocyte progenitor (TIP) cells produce ionocytes as a default lineage. However, Type 2 and Type 17 cytokines divert TIP cell lineage in vitro, resulting in the production of mature tuft cells at the expense of ionocyte differentiation. Our dataset thus provides an updated understanding of airway rare cell composition, and further suggests that clinically relevant cytokines may skew the composition of disease-relevant rare cells.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: Aviv Regev is a founder and equity holder of Celsius Therapeutics, an equity holder in Immunitas Therapeutics, and until August 31, 2020, was a SAB member of Syros Pharmaceuticals, Neogene Therapeutics, Asimov and Thermo Fisher Scientific. From August 1, 2020, Aviv Regev has been an employee of Genentech and has equity in Roche. Avinash Waghray is an employee of Immunology and Inflammation Research Therapeutic Area, Sanofi, Cambridge, MA. Srinivas Vinod Saladi is a co-founder of Reinwik Inc. Brian Lin is a consultant for Rhino Therapeutics and a co-founder of Cellsor. The remaining authors declare no competing interests.
Figures




References
-
- Braga, F. A. V. et al. A cellular census of human lungs identifies novel cell states in health and in asthma. Nat. Med. 10.1038/s41591-019-0468-5 (2019). - PubMed
MeSH terms
Substances
Grants and funding
- TSANKO20G0/Cystic Fibrosis Foundation (CF Foundation)
- 003338L121/Cystic Fibrosis Foundation (CF Foundation)
- 5T32HL116275/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R01 HL164563/HL/NHLBI NIH HHS/United States
- R56 HL157632/HL/NHLBI NIH HHS/United States
- R21 HL156124/HL/NHLBI NIH HHS/United States
- R56HL157632/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- 5R01HL164563/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R01 HL015722/HL/NHLBI NIH HHS/United States
- U24HL148865-01/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R21HL156124/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- U24 HL148865/HL/NHLBI NIH HHS/United States
- R21 HL161760/HL/NHLBI NIH HHS/United States
- U24HL148865-04/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- 5R01HL15722/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- Lin19F0/Cystic Fibrosis Foundation (CF Foundation)
- T32 HL116275/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
