

Brown adipose tissue secretes OLFM4 to coordinate sensory and sympathetic innervation via Schwann cells
- PMID: 40467585
- PMCID: PMC12137647
- DOI: 10.1038/s41467-025-60474-1
Brown adipose tissue secretes OLFM4 to coordinate sensory and sympathetic innervation via Schwann cells
Abstract
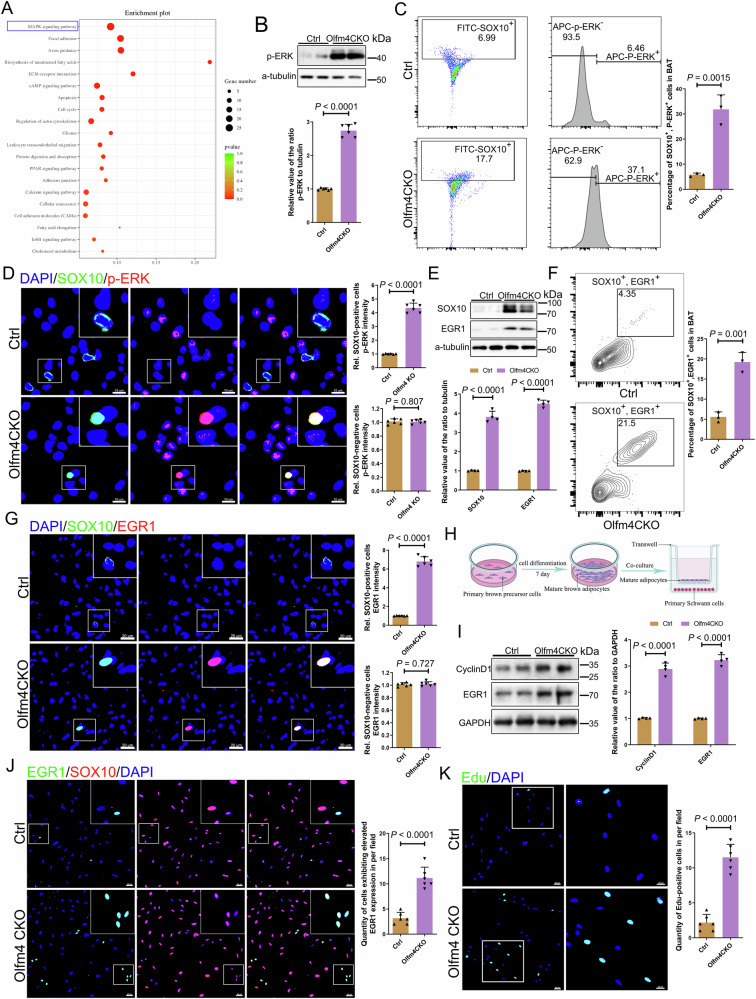
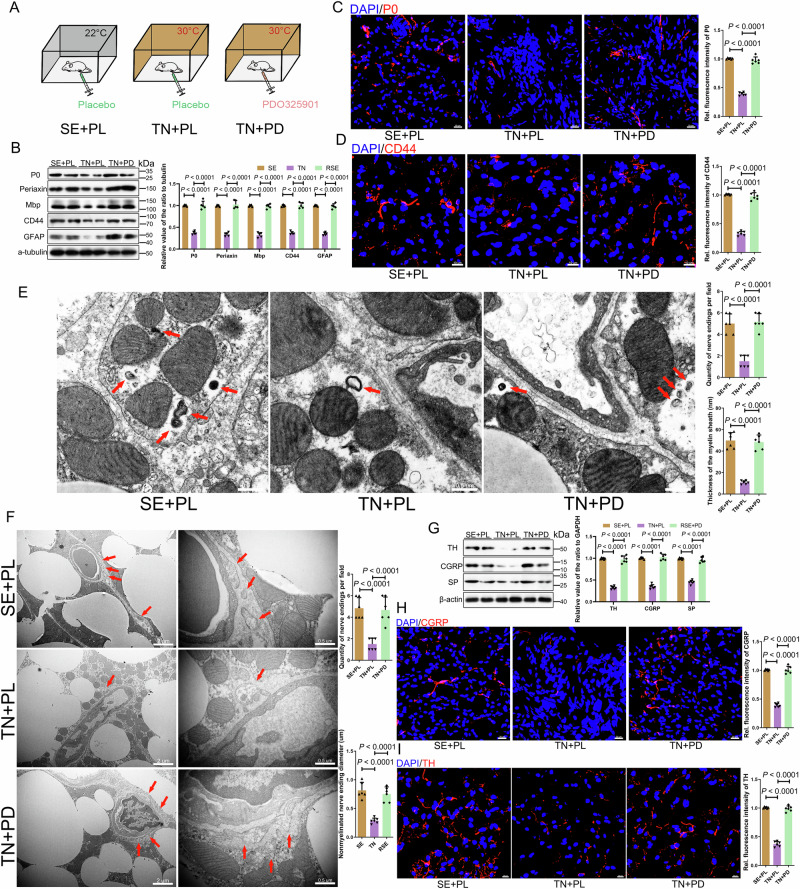
Non-shivering thermogenesis of brown adipose tissue (BAT) is tightly controlled by neural innervation. However, the underlying mechanism remains unclear. Here, we reveal that BAT regulates its own thermoadaptive innervation by crosstalk with Schwann cells (SCs). Loss of Olfm4 (encoding Olfactomedin-4), a risk gene in human obesity, causes BAT dysfunction and reduces whole-body thermogenesis, predisposing to obesity in mice. Mechanistically, BAT-derived OLFM4 traps Noggin, an endogenous inhibitor of BMPs, liberating BMP7-BMPR1B signaling to promote SC differentiation. Conversely, Olfm4 loss reduced BMP7 signaling in mature SCs, leading to MEK/ERK-dependent dedifferentiation and dysfunction, ultimately impairing both sensory and sympathetic innervation. Thermoneutrality exposure reduces Olfm4 expression in BAT, resulting in a similar phenotype. MEK/ERK inhibition, ERK1 depletion, or cold exposure reverses this SC dedifferentiation, enhancing resistance to obesity. These findings suggest that this neurotrophic BAT-SC crosstalk controls thermoadaptive BAT innervation. Reactivating OLFM4 signaling may be a promising therapeutic strategy for obesity and related metabolic diseases.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures






Similar articles
-
Smooth muscle cell-derived Cxcl12 directs macrophage accrual and sympathetic innervation to control thermogenic adipose tissue.Cell Rep. 2024 May 28;43(5):114169. doi: 10.1016/j.celrep.2024.114169. Epub 2024 Apr 27. Cell Rep. 2024. PMID: 38678562 Free PMC article.
-
cAMP-inducible coactivator CRTC3 attenuates brown adipose tissue thermogenesis.Proc Natl Acad Sci U S A. 2018 Jun 5;115(23):E5289-E5297. doi: 10.1073/pnas.1805257115. Epub 2018 May 21. Proc Natl Acad Sci U S A. 2018. PMID: 29784793 Free PMC article.
-
Intact innervation is essential for diet-induced recruitment of brown adipose tissue.Am J Physiol Endocrinol Metab. 2019 Mar 1;316(3):E487-E503. doi: 10.1152/ajpendo.00443.2018. Epub 2018 Dec 21. Am J Physiol Endocrinol Metab. 2019. PMID: 30576247 Free PMC article.
-
Sympathetic and sensory innervation of brown adipose tissue.Int J Obes (Lond). 2010 Oct;34 Suppl 1(0 1):S36-42. doi: 10.1038/ijo.2010.182. Int J Obes (Lond). 2010. PMID: 20935665 Free PMC article. Review.
-
Endogenous ways to stimulate brown adipose tissue in humans.Ann Med. 2015 Mar;47(2):123-32. doi: 10.3109/07853890.2013.874663. Epub 2014 Feb 13. Ann Med. 2015. PMID: 24521443 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
