

Androgens inhibit protective CD8+ T cell responses against pre-erythrocytic malaria parasites in mice
- PMID: 40467616
- PMCID: PMC12137616
- DOI: 10.1038/s41467-025-60193-7
Androgens inhibit protective CD8+ T cell responses against pre-erythrocytic malaria parasites in mice
Abstract
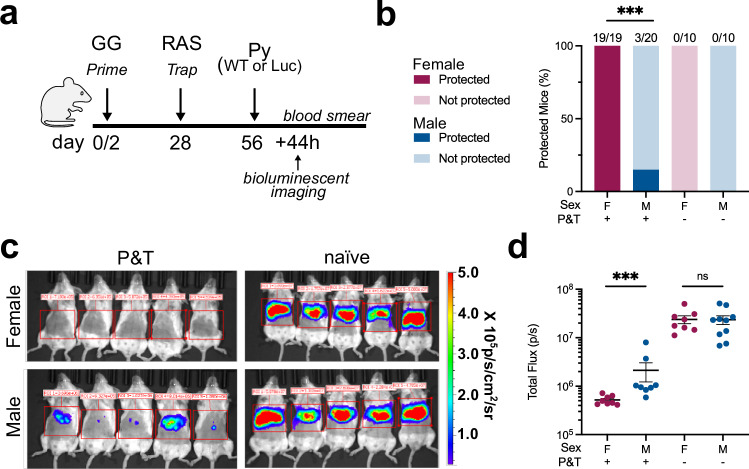
Attenuated whole organism vaccines targeting the malaria liver stage reliably confer sterile immunity. These vaccines completely protect female mice from infection, but protection in male mice remains unproven. We discover that male mice vaccinated with prime-and-trap, a whole organism-based vaccine strategy, exhibit poorer protection against Plasmodium sporozoite challenge than females. We investigate this sex difference, and identify vaccinated males have fewer hepatic memory CD8+ T cells than females when scaling for liver biomass, and reduced inflammatory responses post-vaccination. Surgical hormone manipulation clarifies that the presence of testicular hormones hinders protection in male mice. The presence of androgens does not affect memory CD8+ T cell quantity nor quality, but reduces recruitment of CD8+ T cells in male liver tissues via a restricted inflammatory response. Here, we show both males and females form functional memory responses following prime-and-trap vaccination, but the presence of androgens during sporozoite challenge impair protection in male mice.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: C.J.D., N.S., F.N.W., A.S.P., E.D.L., K.B., E.N.C, A.C.K., A.M.S, C.S. and M.J.S. declare no competing interests. S.C.M. filed a patent application on selected aspects of the prime-and-trap concept through the University of Washington. S.C.M. has equity in a startup company (Sound Vaccines, Inc.) that is negotiating with the University of Washington for rights to this intellectual property. The relationship between the authors and Sound Vaccines, Inc., has been reviewed by the University of Washington and complies with all University and State of Washington policies on such activities. K.A.D. is a paid contractor for Seattle Biosoftware, which had no input or association with this study.
Figures






References
-
- World Malaria Report 2024. (World Health Organization, Geneva, 2024).
-
- Cowman, A. F., Healer, J., Marapana, D. & Marsh, K. Malaria: biology and disease. Cell167, 610–624 (2016). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
