

This is a preprint.
Calcium channel-coupled transcription factors facilitate direct nuclear signaling
- PMID: 40470204
- PMCID: PMC12136203
- DOI: 10.21203/rs.3.rs-6180510/v1
Calcium channel-coupled transcription factors facilitate direct nuclear signaling
Abstract
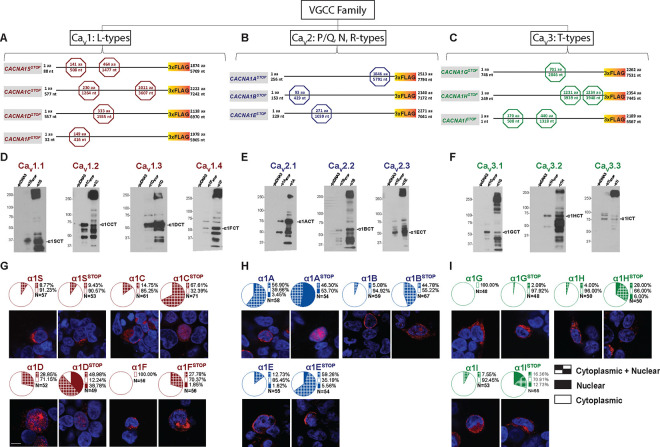
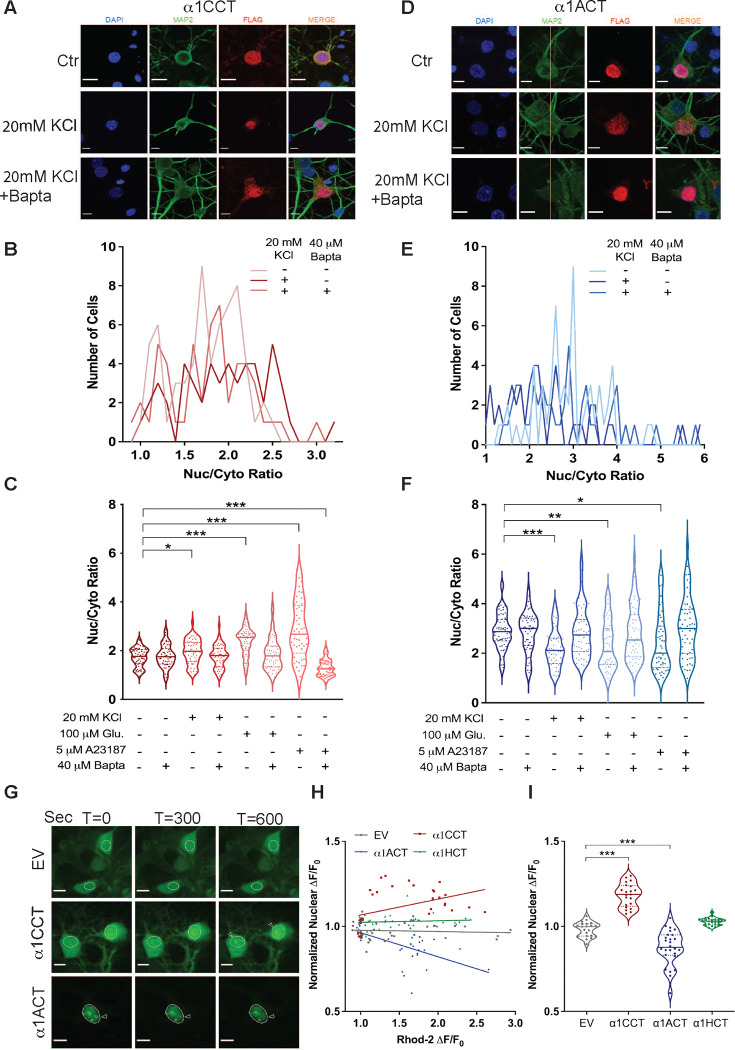
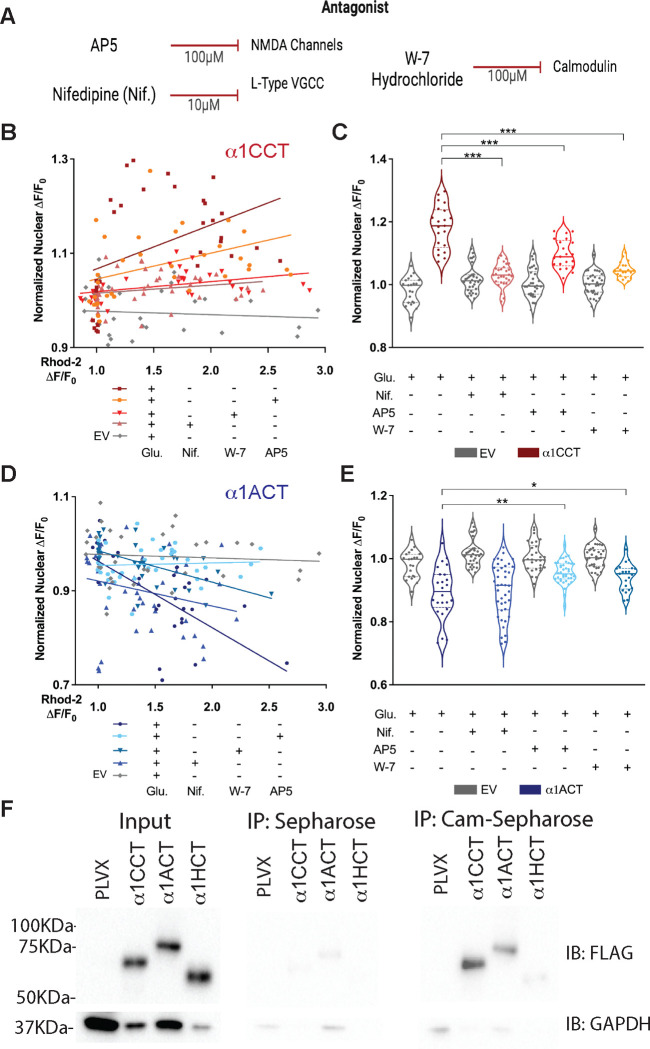
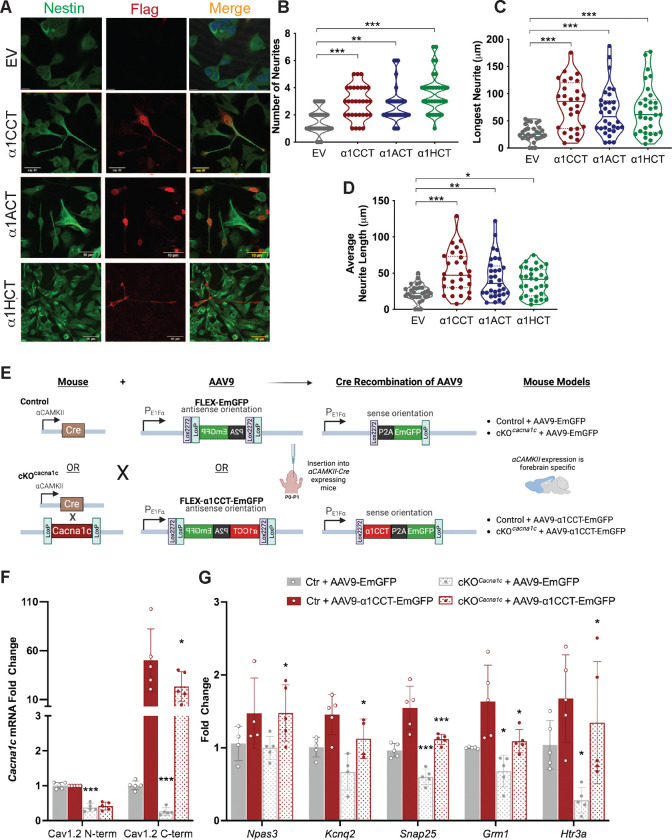
VGCCs play crucial roles within the CNS, in maintaining cell excitability, enabling activity-dependent neuronal development, and forming long-term memory by regulating Ca2+ influx. The intracellular carboxyl-terminal domains of VGCC α1 subunits help regulate VGCC function. Emerging evidence suggests that some VGCC C-termini have functions independent of channel gating and exist as stable proteins. Here, we demonstrate that all VGCC gene family members express bicistronic mRNA transcripts that produce functionally distinct C-terminal proteins (CTPs) in tandem with full-length VGCC α1 subunits. Two of these CTPs, α1CCT and α1ACT, cycle to and from the nucleus in a Ca2+- and calmodulin-dependent fashion. α1CCT, α1ACT, and α1HCT regulate chromatin accessibility and/or bind directly to genes, regulating gene networks involved in neuronal differentiation and synaptic function in a Ca2+-dependent manner. This study elucidates a conserved process of coordinated protein expression within the VGCC family, coupling the channel function with VGCC C-terminal transcription factors.
Keywords: C-terminal proteins; CACNA1A; CACNA1C; CACNA1H; Ca2+ signaling; VGCC; Voltage-gated Ca2+ channels; bicistronic cellular genes; neuronal differentiation; synaptic function; α1ACT; α1CCT; α1HCT.
Conflict of interest statement
Author Declaration of Interests The authors declare no competing interests.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
