

L-serine promotes pro-carcinogenic effects of colibactin-producing E. coli
- PMID: 40471070
- PMCID: PMC12143686
- DOI: 10.1080/19490976.2025.2515480
L-serine promotes pro-carcinogenic effects of colibactin-producing E. coli
Abstract
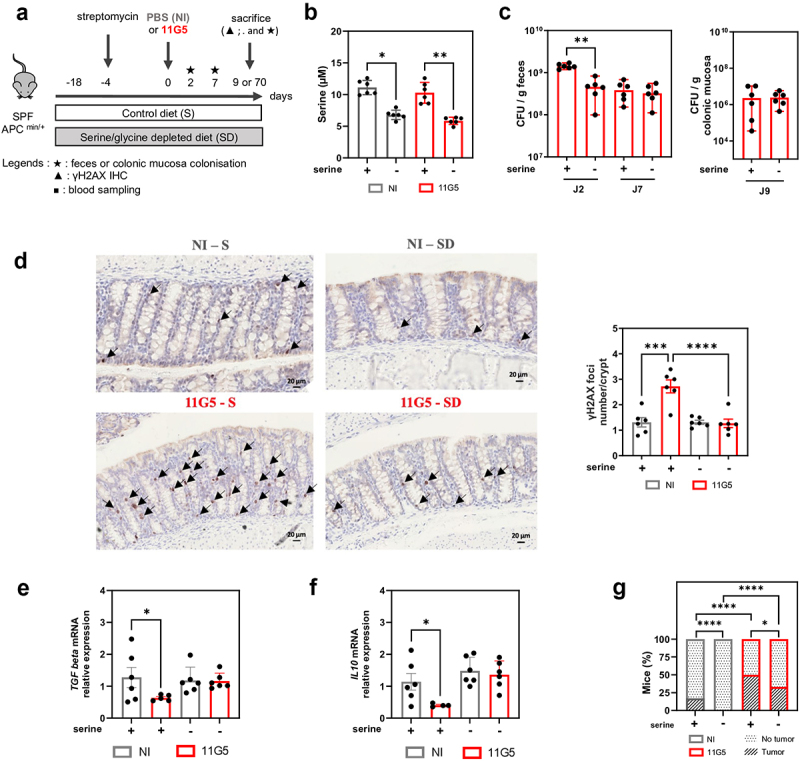
Colonic tissues are abnormally colonized by colibactin-producing Escherichia coli (CoPEC) in colorectal cancer (CRC) patients. CoPECs have been shown to promote colorectal carcinogenesis in several pre-clinical CRC mouse models. Here, we report that CoPEC reprograms the metabolism of colonic epithelial cells in a colibactin-dependent manner, leading to a Warburg-like effect, altered redox homeostasis, and disrupted amino acid metabolism. Among these metabolic modifications, we observed a significant decrease in both extracellular and intracellular serine levels. We found that CoPEC activates the L-serine-utilization operon during gut colonization, maximizing its competitive fitness advantage over a commensal strain. Moreover, an L-serine-depleted diet induces an early and transient decrease in CoPEC colonization of mice gut, associated with decrease of both DNA damages and tumor development. Finally, deletion of the bacterial tdcA gene involved in L-serine operon utilization reduces the competitive fitness of CoPEC, the in vitro adhesion and persistence within the epithelial cells and leads in CRC animal models to reduced carcinogenic activity of the pathobiont. This work highlights the interplay between intestinal microbiota factors, such as CoPEC, and nutritional factors, such as L-serine, in colorectal carcinogenesis.
Keywords: CoPEC; Colorectal cancer; L-serine; colibactin producing E. coli; pks island.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures





References
-
- Cancer (IARC) TIA for R on. Global Cancer Observatory [Internet]. [cited 2024 Jan 11]. https://gco.iarc.fr/.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous