

A role of arginase-1-expressing myeloid cells in cachexia
- PMID: 40474277
- PMCID: PMC12142917
- DOI: 10.1186/s40170-025-00396-0
A role of arginase-1-expressing myeloid cells in cachexia
Abstract
Background: Despite decades of efforts to find successful treatment approaches, cachexia remains a major unmet medical need. This condition, that affects patients with diverse underlying conditions, is characterized by severe muscle loss and is associated with reduced quality of life and limited survival. Search for underlying mechanisms that may guide cachexia treatment has mainly evolved around potential atrophy-inducing roles of inflammatory mediators, and in cancer patients, tumor-derived factors. Recently, a new paradigm emerged as it is becoming evident that specific immune cells inhabit atrophic muscle tissue. Arginase 1 (Arg1) expression is characteristic of these immune cells. Studies of potential contributions of these immune cells to loss of muscle mass and function is in its infancy, and the contribution of ARG1 to these processes remains elusive.
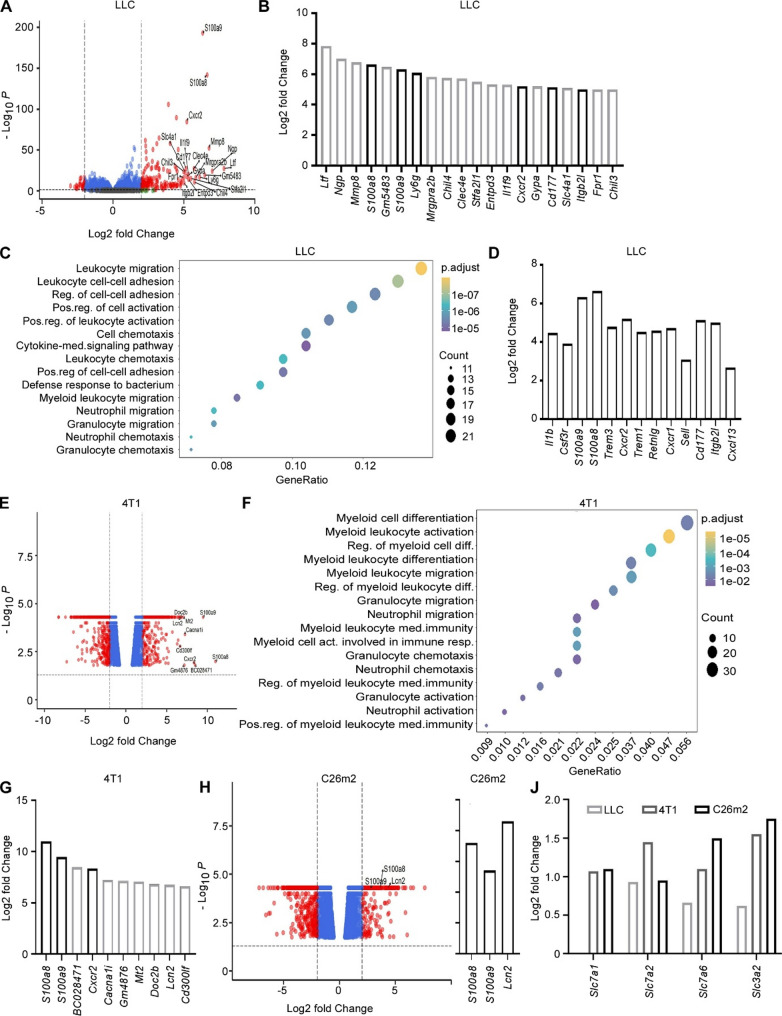
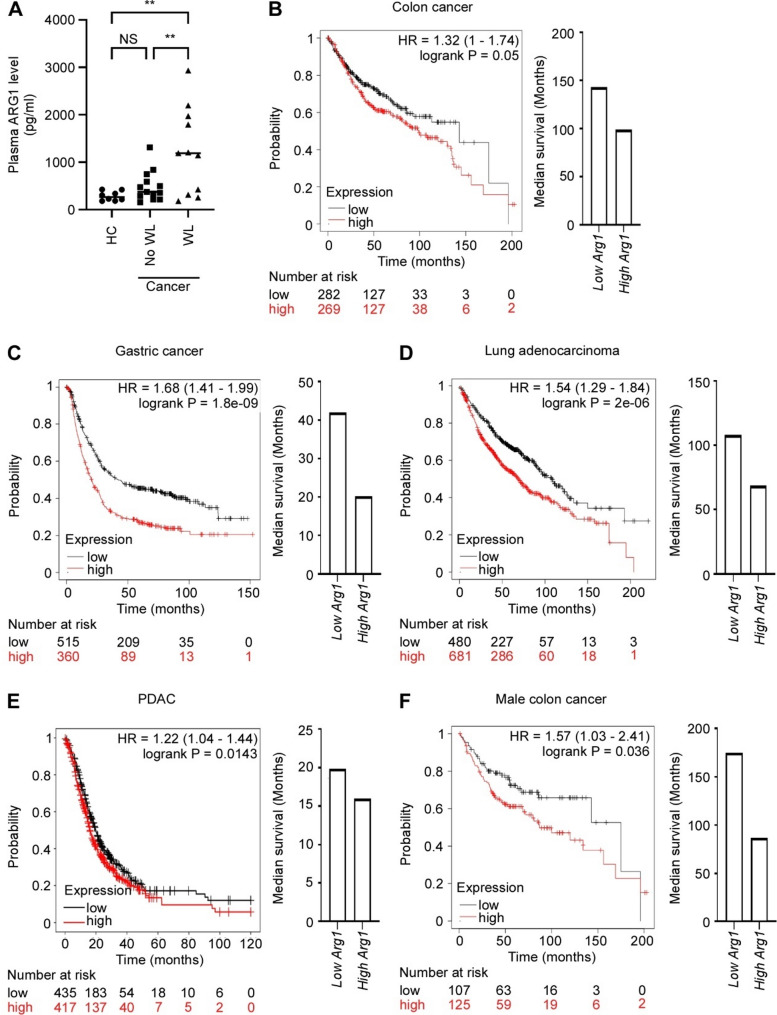
Methods: Analyses of RNA sequencing data from murine cachexia models and comprehensive, unbiased open approach proteomics analyses of skeletal myotubes was performed. In vitro techniques were employed to evaluate mitochondrial function and capacity in skeletal muscle cells and cardiomyocytes. Functional bioassays were used to measure autophagy activity. ARG1 level in patients' plasma was evaluated using ELISA, and the association between ARG1 level and patient survival, across multiple types of cancer, was examined using the online database Kaplan-Meier plotter.
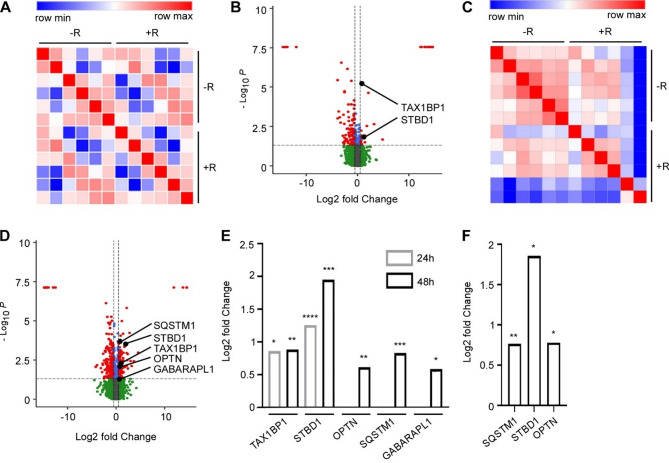
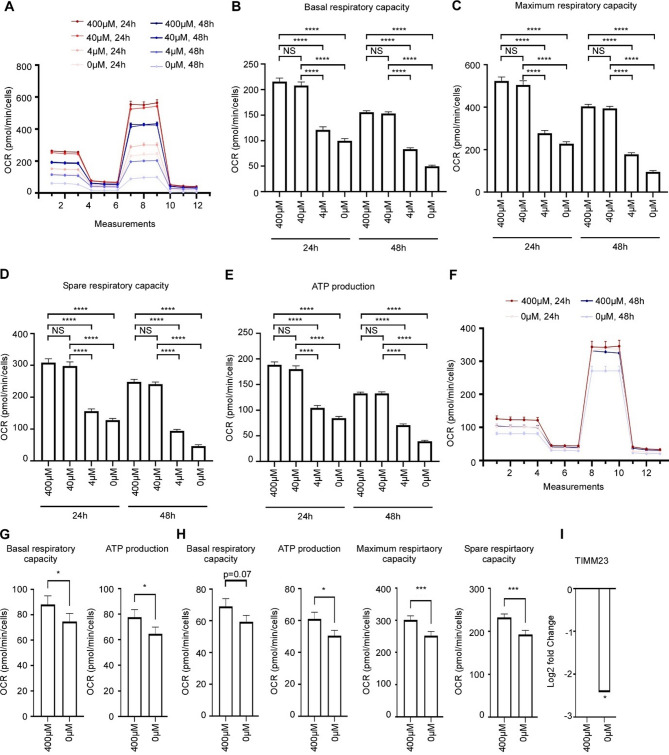
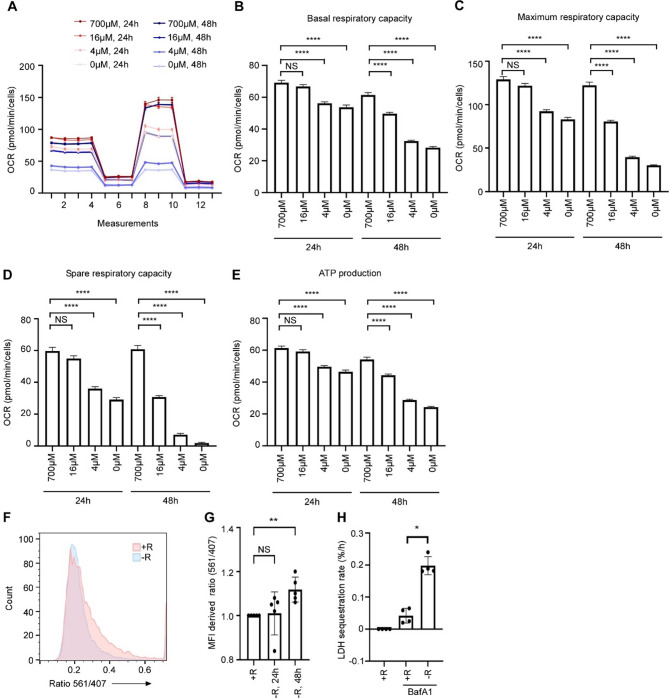
Results: In line with arginine-degrading activity of ARG1, we found signs of arginine restriction in atrophic muscles. In response to arginine restriction, mitochondrial functions and ATP generation was severely compromised in both skeletal muscle cells and in cardiomyocytes. In skeletal muscle cells, arginine restriction enhanced the expression of autophagic proteins, suggesting autophagic degradation of cellular content. Reduction in mitochondria marker TIMM23 supports selective autophagic degradation of mitochondria (mitophagy). In arginine starved cardiomyocytes, mitochondrial dysfunction is accompanied by both increased bulk autophagy and mitophagy. In cancer patients, we found an association between ARG1 expression and accelerated weight loss and reduced survival, further supporting a role of ARG1-producing cells in cachexia pathogenesis.
Conclusion: Together, our findings point to a mechanism for cachexia which depends on expansion of ARG1-expressing myeloid cells, local restriction of arginine, loss of mitochondrial capacity and induced catabolism in skeletal muscle cells and in the heart.
Keywords: ARG1; Arginine; Autophagy; Cachexia; Cancer; Mitochondria; Mitophagy; Muscle; Myeloid-derived suppressor cell; Neutrophil.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethical approval and consent to participate: Experiments involving patient material were performed in accordance with the approval from the local ethical committee (Lothian Regional Ethics Committee (REC), Scotland, 06/S1103/75). Written informed consent was received from participants prior to inclusion in the study and the study was performed in conformity with the declaration of Helsinki. Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures
References
-
- Vagnildhaug OM, Balstad TR, Almberg SS, Brunelli C, Knudsen AK, Kaasa S, et al. A cross-sectional study examining the prevalence of cachexia and areas of unmet need in patients with cancer. Supportive Care Cancer: Official J Multinational Association Supportive Care Cancer. 2018;26(6):1871–80. - PubMed
-
- Tesi RJ. MDSC; the most important cell you have never heard of. Trends Pharmacol Sci. 2019;40(1):4–7. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
