

This is a preprint.
Febrile temperature activates the innate immune response by promoting aberrant influenza A virus RNA synthesis
- PMID: 40475563
- PMCID: PMC12139806
- DOI: 10.1101/2025.05.19.654939
Febrile temperature activates the innate immune response by promoting aberrant influenza A virus RNA synthesis
Update in
-
Febrile temperature activates the innate immune response by promoting aberrant influenza A virus RNA synthesis.Sci Adv. 2026 Jan 2;12(1):eaeb2700. doi: 10.1126/sciadv.aeb2700. Epub 2026 Jan 2. Sci Adv. 2026. PMID: 41481744 Free PMC article.
Abstract
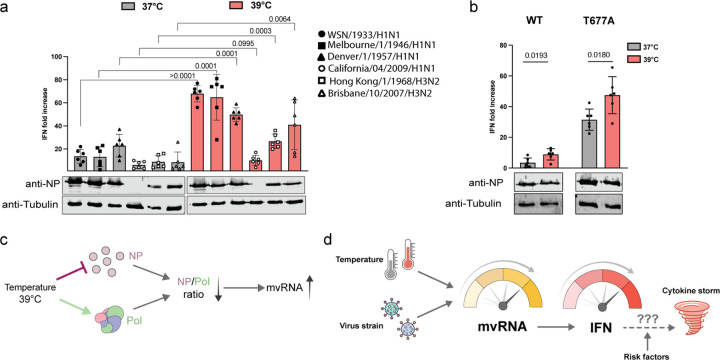
Fever during influenza A virus (IAV) infection is triggered by the innate immune response. Various factors contribute to this response, including IAV mini viral RNAs (mvRNA), which trigger RIG-I signaling when their replication and transcription are dysregulated by template loops (t-loop). It is presently not well understood whether the fever response to IAV infection impacts subsequent viral replication and innate immune activation. Here we show that IAV infection at temperatures that simulate fever leads to increased antiviral signaling in H1N1 and H3N2 infections. Mathematical modeling and experimental analyses reveal that differential IAV nucleoprotein and RNA polymerase production increase mvRNA and interferon production. Moreover, at the higher infection temperature, mvRNAs with dysregulating t-loops contribute most to the innate immune activation. We propose that fever during IAV infection can establish a positive feedback loop in which elevated aberrant RNA synthesis and innate immune activation can contribute to the dysregulation of cytokine production.
Keywords: RIG-I; RNA polymerase; capped cRNA; fever; influenza A virus; interferon; mini viral RNA; mvRNA; temperature; template loop.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures
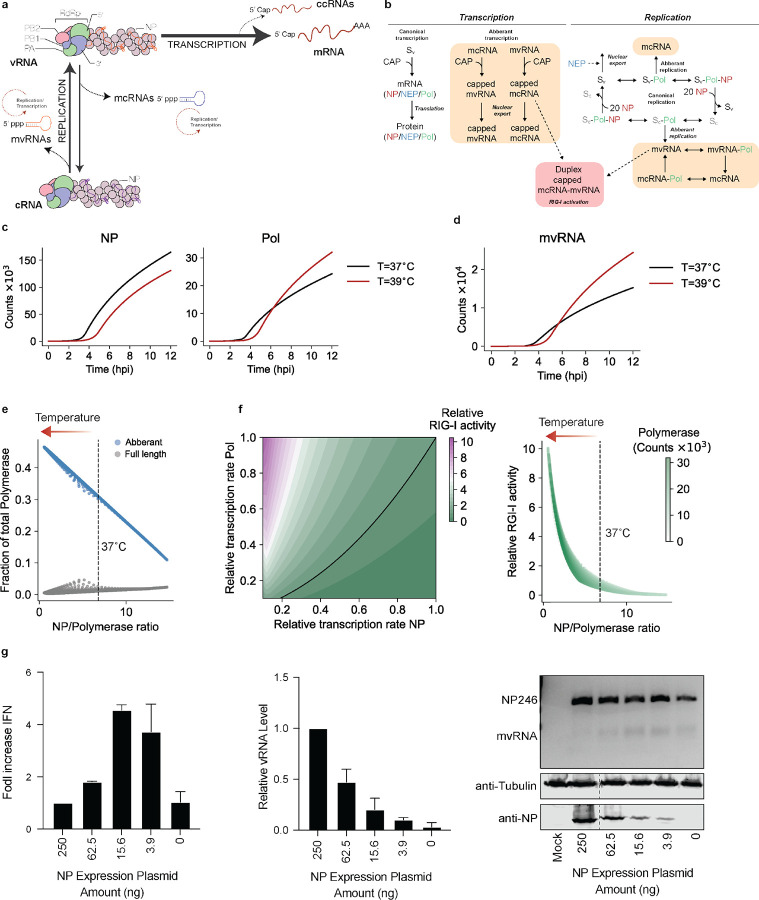
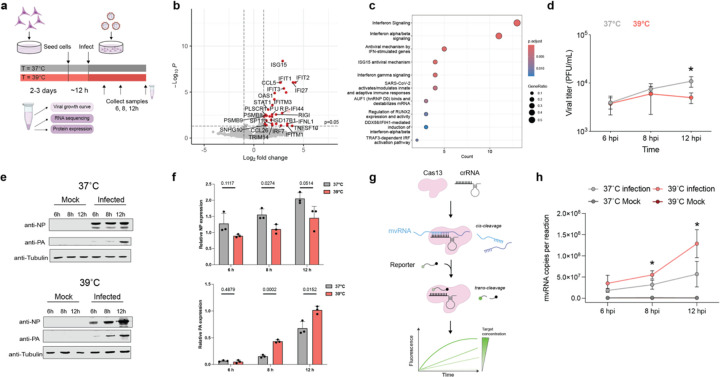
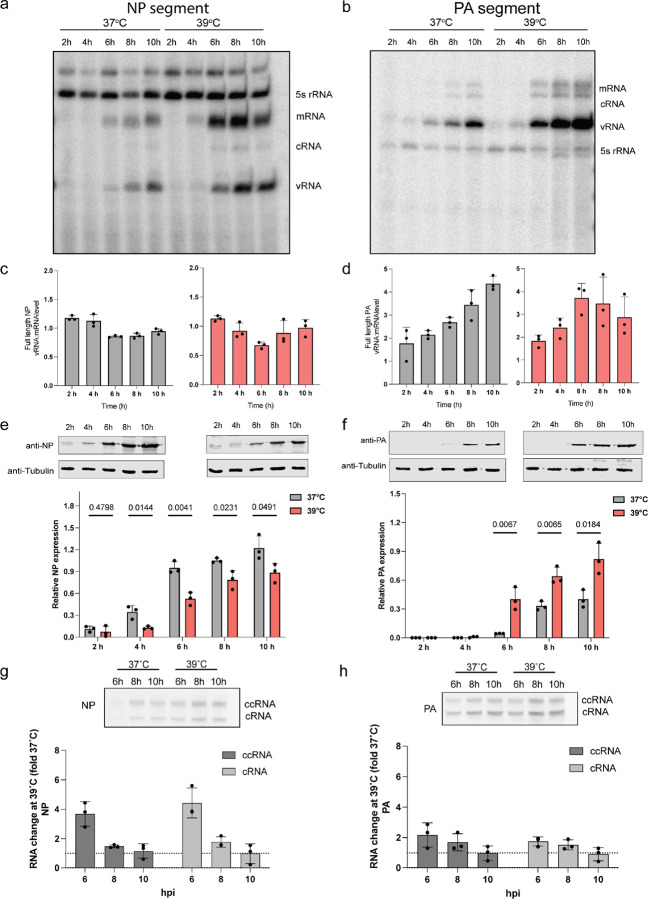
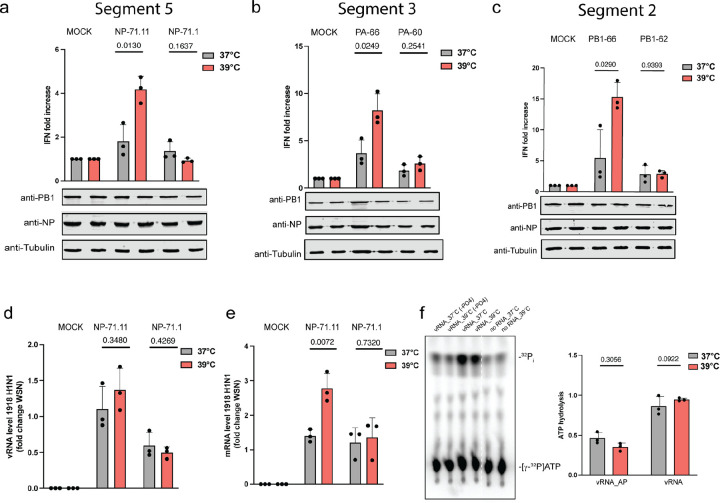
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources