

Rapid and reversible fluorescent probe enables repeated snapshot imaging of AMPA receptors during synaptic plasticity
- PMID: 40479050
- PMCID: PMC12143348
- DOI: 10.1126/sciadv.adt6683
Rapid and reversible fluorescent probe enables repeated snapshot imaging of AMPA receptors during synaptic plasticity
Abstract
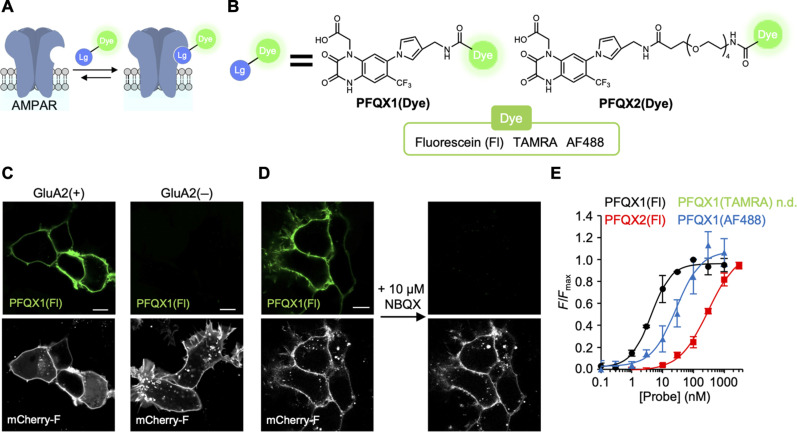
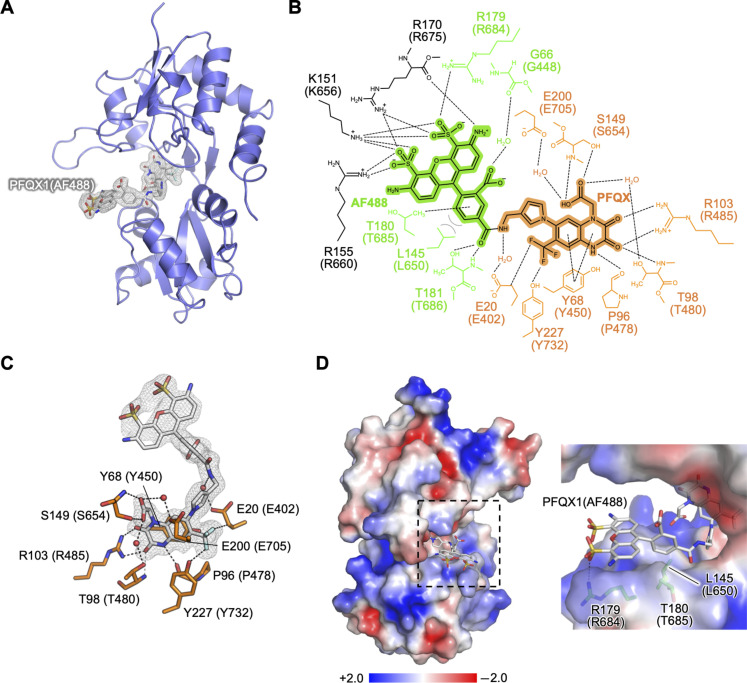
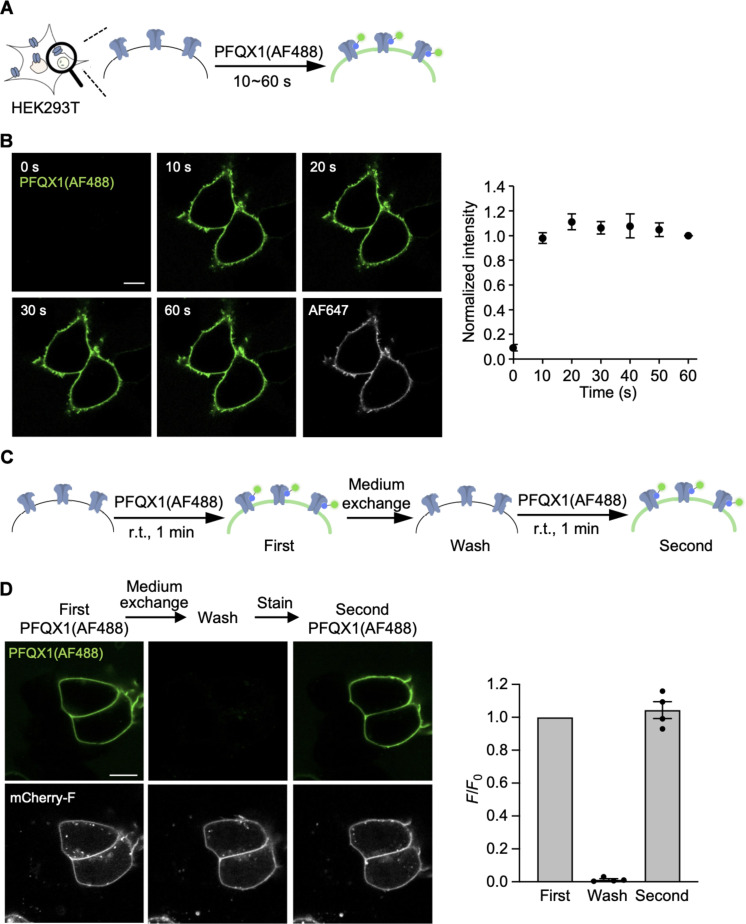
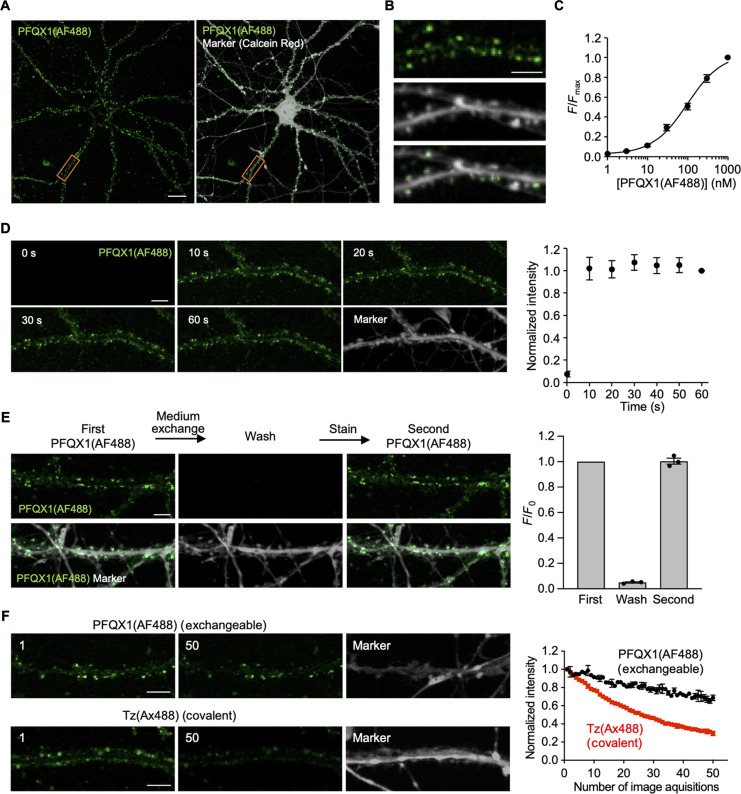
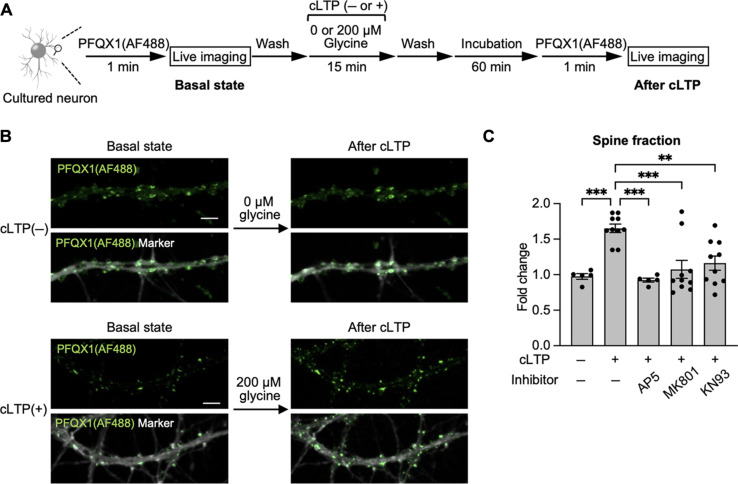
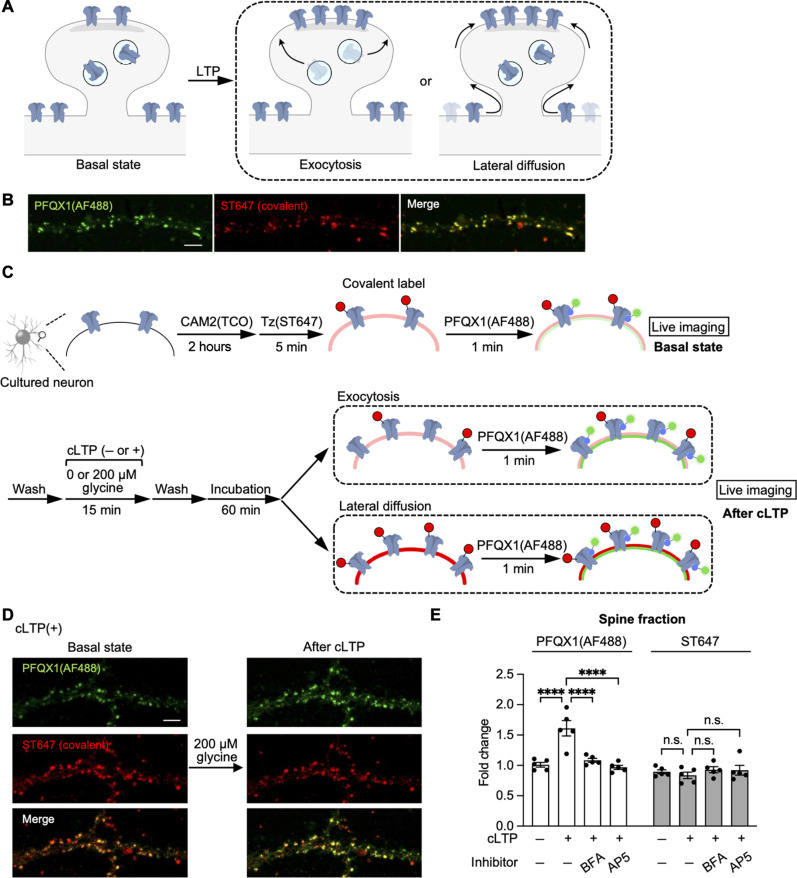
The subcellular localization of neurotransmitter receptors is strictly regulated in neurons. Changes in the trafficking of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)-type glutamate receptors (AMPARs) play an essential role in synaptic plasticity, which is the cellular basis of learning and memory. To explore receptor trafficking, genetically encoded approaches (e.g., the fusion of fluorescent proteins to receptors) are often used. However, concerns remain that genetic approaches cannot fully reproduce the receptor functions that are inherent to neurons. Herein, we report on PFQX1(AF488), a fluorescent probe for the visualization of cell-surface AMPARs without any genetic manipulation to neurons. The rapid and reversible staining features of this probe enabled snapshot imaging, which showed the accumulation of native AMPARs in dendritic spines during synaptic plasticity. Moreover, the mechanism of this synaptic accumulation, for which genetically encoded approaches have given controversial results, was revealed by integrating two chemical methods: PFQX1(AF488) and covalent chemical labeling.
Figures
Similar articles
-
Spatiotemporal modelling reveals geometric dependence of AMPAR dynamics on dendritic spine morphology.J Physiol. 2023 Aug;601(15):3329-3350. doi: 10.1113/JP283407. Epub 2022 Nov 23. J Physiol. 2023. PMID: 36326020
-
Long-term potentiation in isolated dendritic spines.PLoS One. 2009 Jun 23;4(6):e6021. doi: 10.1371/journal.pone.0006021. PLoS One. 2009. PMID: 19547754 Free PMC article.
-
Crucial Role of Postsynaptic Syntaxin 4 in Mediating Basal Neurotransmission and Synaptic Plasticity in Hippocampal CA1 Neurons.Cell Rep. 2018 Jun 5;23(10):2955-2966. doi: 10.1016/j.celrep.2018.05.026. Cell Rep. 2018. PMID: 29874582
-
Dynamic imaging of AMPA receptor trafficking in vitro and in vivo.Curr Opin Neurobiol. 2017 Aug;45:51-58. doi: 10.1016/j.conb.2017.03.008. Epub 2017 Apr 12. Curr Opin Neurobiol. 2017. PMID: 28411409 Free PMC article. Review.
-
Regulation of AMPAR trafficking in synaptic plasticity by BDNF and the impact of neurodegenerative disease.J Neurosci Res. 2022 Apr;100(4):979-991. doi: 10.1002/jnr.25022. Epub 2022 Feb 7. J Neurosci Res. 2022. PMID: 35128708 Review.
References
-
- Bliss T. V., Collingridge G. L., A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 361, 31–39 (1993). - PubMed
-
- Martin S. J., Grimwood P. D., Morris R. G., Synaptic plasticity and memory: An evaluation of the hypothesis. Annu. Rev. Neurosci. 23, 649–711 (2000). - PubMed
-
- Citri A., Malenka R. C., Synaptic plasticity: Multiple forms, functions, and mechanisms. Neuropsychopharmacology 33, 18–41 (2008). - PubMed
-
- Bredt D. S., Nicoll R. A., AMPA receptor trafficking at excitatory synapses. Neuron 40, 361–379 (2003). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
