

Targeting PERP promotes anti-tumor immunity in HNSCC by regulating tumor immune microenvironment and metabolic homeostasis
- PMID: 40483487
- PMCID: PMC12144827
- DOI: 10.1186/s12943-025-02360-4
Targeting PERP promotes anti-tumor immunity in HNSCC by regulating tumor immune microenvironment and metabolic homeostasis
Abstract
Background: PERP may have the potential to function as an oncogene. However, the precise function, prognostic value, and predictive significance remain shrouded in ambiguity.
Methods: We conducted an in-depth analysis using pan-cancer RNA sequencing data and various online web tools to investigate the correlation between PERP and crucial clinical outcomes such as prognosis, tumor microenvironment, and tumor metabolism. In addition, we explored the tumor-promoting role of PERP and its potential mechanisms through models such as immunofluorescence staining, flow cytometry, cell proliferation assays, wound healing assays, cell migration assays, mass spectrometry analysis and isotope tracing. Further in vivo models confirmed the functional consistency of PERP across pan-cancer. Finally, we analyzed the potential of PERP as a predictive factor for immunotherapy sensitivity in a clinical cohort.
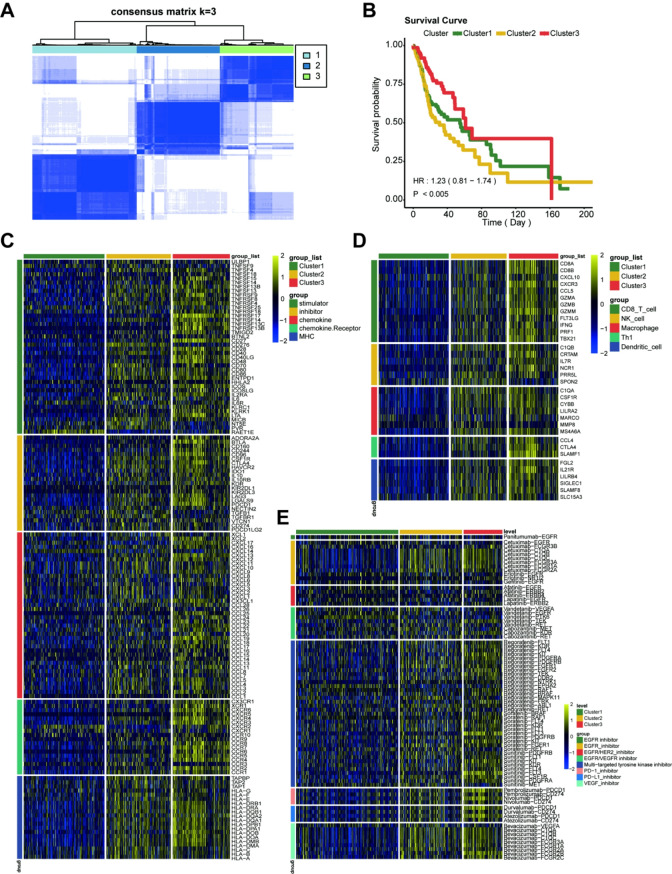
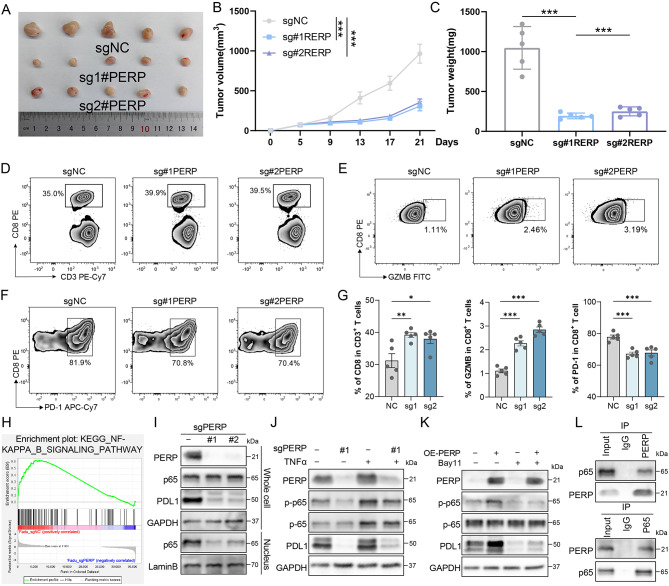
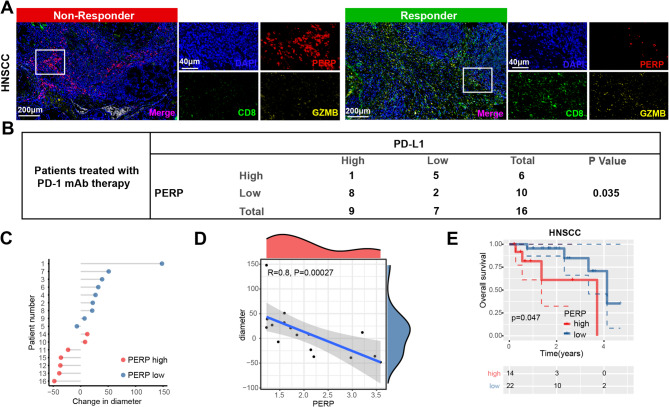
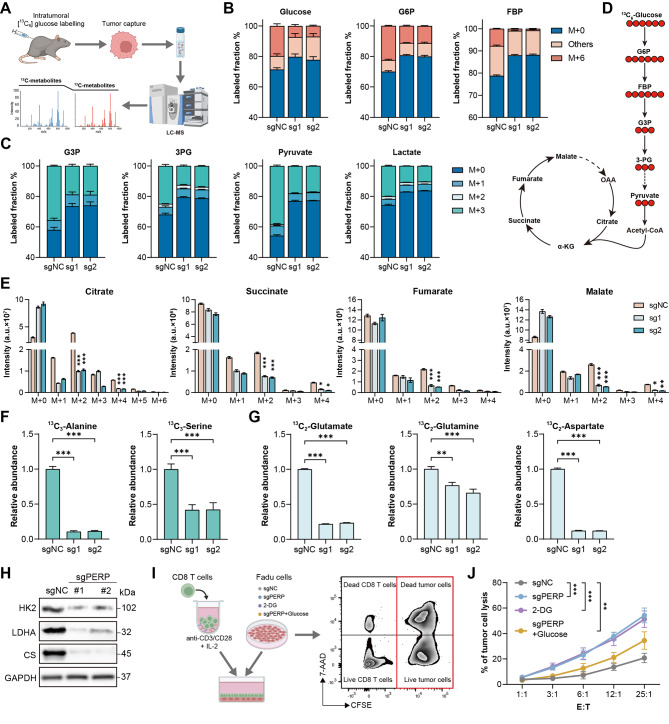
Results: PERP exhibits elevated expression in the majority of cancer types and impedes immune cell infiltration as well as immune checkpoint reactivity in pan-cancer. We confirmed that PERP can promote tumor progression by tumor cell proliferation, scratch and transwell experiments. Meanwhile, the absence of PERP restricts the flux of 13C6-glucose into glycolysis and the tricarboxylic acid (TCA) cycle. Importantly, the deficiency of PERP enhances the in vivo anti-tumor efficacy of PD1 monoclonal antibodies. In addition, low PERP expression is highly correlated with the response of head and neck squamous cell carcinoma (HNSCC) patients to immunotherapy.
Conclusions: PERP represents a promising predictive/diagnostic biomarker and therapeutic target for HNSCC patients.
Keywords: Head and neck squamous cell carcinoma; PD1; PERP; Prognosis; Tumor microenvironment.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: Experiments involving clinical samples were carried out in compliance with the approved protocol of the Clinical Research Ethics Committee of the Xiangya Hospital Central South University (protocol 2023030315). Experiments involving animals were approved by the Institutional Review Committee of Xiangya Hospital, Central South University (protocol XY20240711004). Consent for publication: Not applicable. Competing interests: The authors declare no competing interests.
Figures



References
-
- Kato K, Cho BC, Takahashi M, Okada M, Lin CY, Chin K, Kadowaki S, Ahn MJ, Hamamoto Y, Doki Y, et al. Nivolumab versus chemotherapy in patients with advanced oesophageal squamous cell carcinoma refractory or intolerant to previous chemotherapy (ATTRACTION-3): a multicentre, randomised, open-label, phase 3 trial. Lancet Oncol. 2019;20(11):1506–17. - DOI - PubMed
MeSH terms
Substances
Grants and funding
- 82303717/National Natural Science Foundation of China
- 82272872/National Natural Science Foundation of China
- 2023M743954/China Postdoctoral Science Foundation
- 2023JJ40992/Natural Science Foundation of Hunan Province
- kq 2208393/Changsha Natural Science Foundation
- 82322048/National Outstanding Youth Science Fund Project of National Natural Science Foundation of China
- JQ2023H004/Natural Science Foundation of Heilongjiang Province
- Nn102024-02/Harbin Medical University Cancer Hospital: Nn10 Plan
- CYQN24011/Chun Yan of Heilongjiang Provincial Science and Technology Department supported the planned construction project
LinkOut - more resources
Full Text Sources
Medical
