

C1q reprograms innate immune memory
- PMID: 40486519
- PMCID: PMC12141851
- DOI: 10.3389/fimmu.2025.1515127
C1q reprograms innate immune memory
Abstract
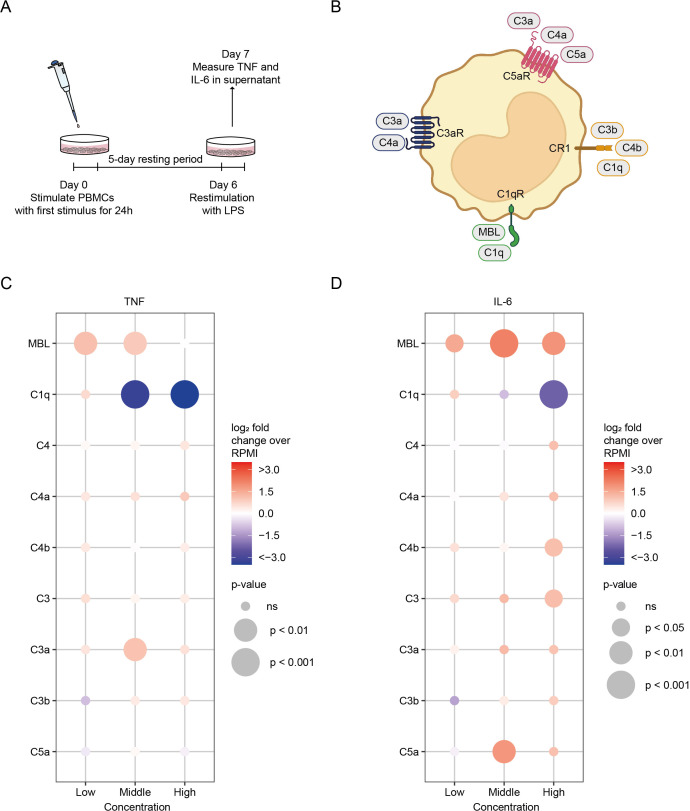
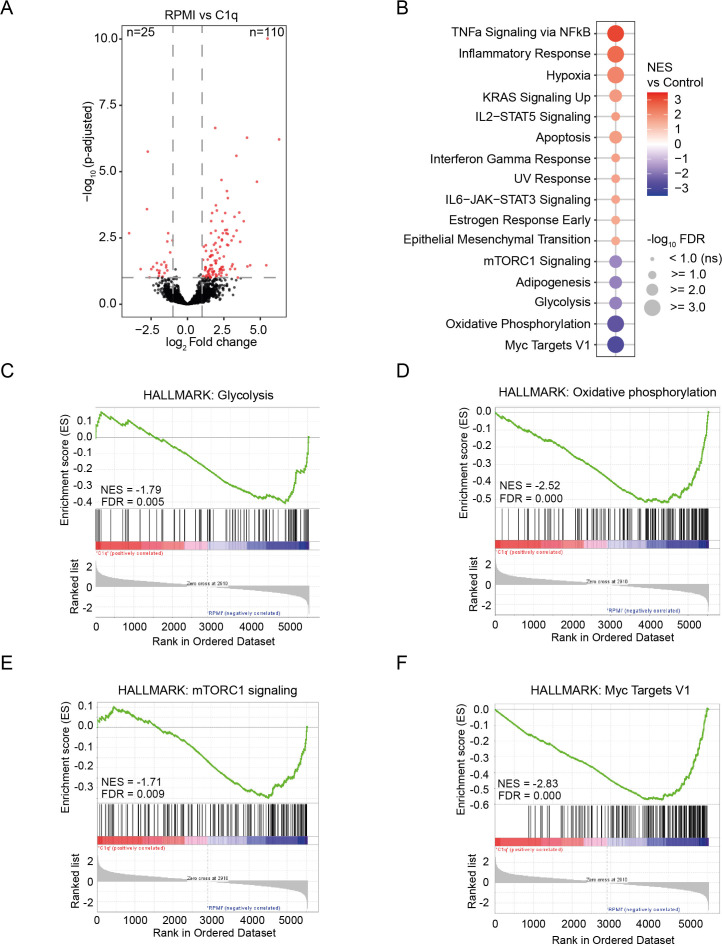
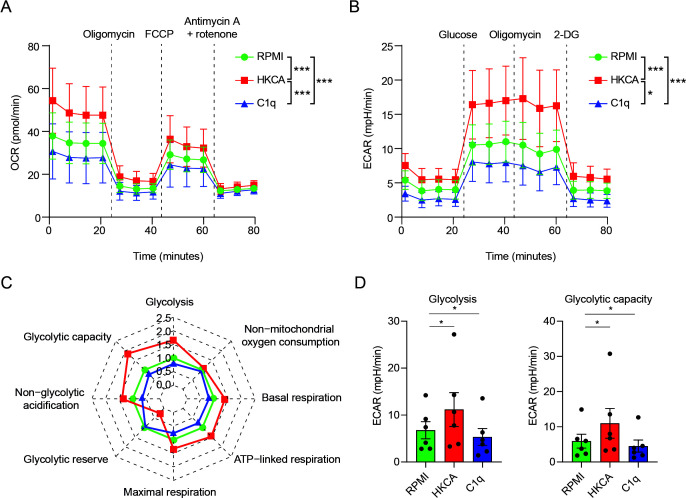
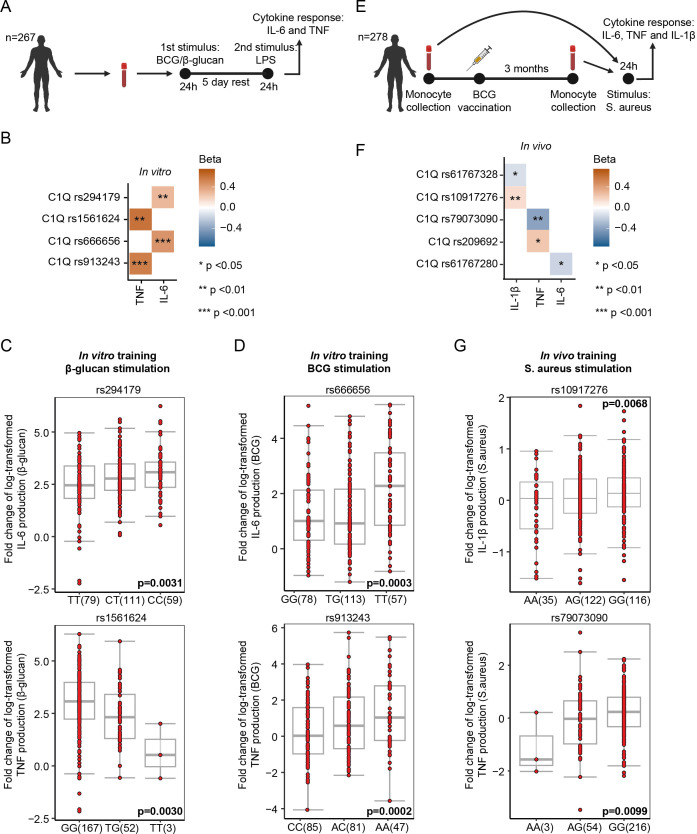
Innate immune memory, also called trained immunity, is a metabolic and epigenetically regulated process that enables innate immune cells to recalibrate their inflammatory reactivity in response to pathogenic or endogenous stimuli. In addition to its function in host defense, trained immunity contributes to diverse immune-mediated diseases. We discovered that complement component 1q (C1q) is an effective modulator of innate immune memory, potently suppressing the responsiveness of myeloid cells. We found that C1q leads to profound reprogramming of myeloid cell metabolism, particularly glycolysis, and exerts control over the transcriptional regulation of important metabolic and inflammatory genes. We corroborate our findings by identifying single-nucleotide polymorphisms close to the C1q gene that are linked to induction of trained immunity by Bacillus Calmette-Guérin (BCG) or beta-glucan in healthy peripheral blood mononuclear cells. Our results reveal an immunomodulatory role for C1q and provide evidence of a molecular interaction between the complement system and innate immune memory. These findings expand our understanding of innate immune memory.
Keywords: C1q; complement; immunometabolism; innate immune memory; tolerance; trained immunity.
Copyright © 2025 Jonkman, Jacobs, Negishi, van Heck, Matzaraki, Martens, Baltissen, Vermeulen, Fayad, Teunissen, Mulder, Hilbrands, Joosten, Netea, Mhlanga, Rother and Duivenvoorden.
Conflict of interest statement
WM is scientific founder of TTxD and Biotrip. LJ is scientific founder of TTxD, LembaTX and SalvinaTX. MN is scientific founder of TTxD, Biotrip, LembaTX, and SalvinaTX. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
The role of sirtuin 1 on the induction of trained immunity.Cell Immunol. 2021 Aug;366:104393. doi: 10.1016/j.cellimm.2021.104393. Epub 2021 Jun 12. Cell Immunol. 2021. PMID: 34147841
-
Differential induction of innate memory in porcine monocytes by β-glucan or bacillus Calmette-Guerin.Innate Immun. 2021 Aug;27(6):448-460. doi: 10.1177/1753425920951607. Epub 2020 Aug 30. Innate Immun. 2021. PMID: 32862748 Free PMC article.
-
The causes and consequences of trained immunity in myeloid cells.Front Immunol. 2024 Apr 11;15:1365127. doi: 10.3389/fimmu.2024.1365127. eCollection 2024. Front Immunol. 2024. PMID: 38665915 Free PMC article. Review.
-
Seasonal variation in BCG-induced trained immunity.Vaccine. 2024 Oct 24;42(24):126109. doi: 10.1016/j.vaccine.2024.07.010. Epub 2024 Jul 8. Vaccine. 2024. PMID: 38981740
-
Infectious Agents as Stimuli of Trained Innate Immunity.Int J Mol Sci. 2018 Feb 3;19(2):456. doi: 10.3390/ijms19020456. Int J Mol Sci. 2018. PMID: 29401667 Free PMC article. Review.
References
-
- Sim R, Dodds A. Complement. A practical approach. Dodds AW, Sim RB, editors. Oxford: IRL; (1997) p. 1–18.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources