

Targeting SUMOylation in ovarian cancer: Sensitivity, resistance, and the role of MYC
- PMID: 40487451
- PMCID: PMC12145850
- DOI: 10.1016/j.isci.2025.112555
Targeting SUMOylation in ovarian cancer: Sensitivity, resistance, and the role of MYC
Abstract
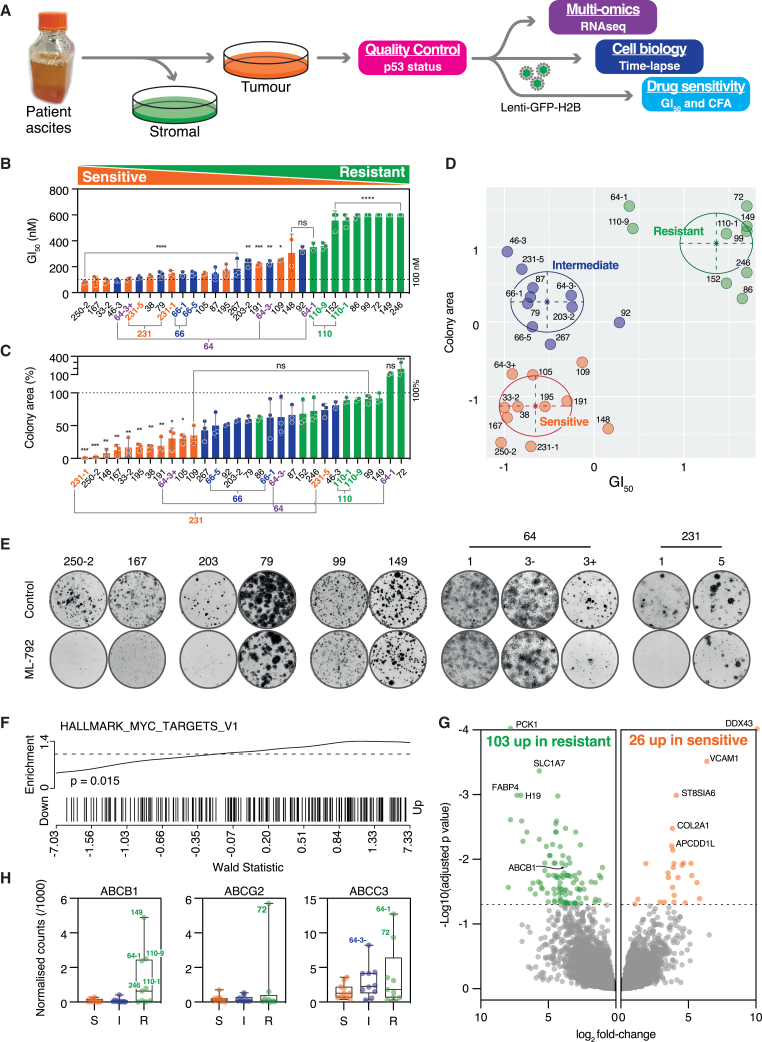
Cells overexpressing MYC depend on SUMOylation for survival and cell division. To assess the therapeutic potential of SUMO inhibition, we screened 30 patient-derived ovarian cancer models (OCMs) with the SUMO-activating enzyme inhibitor ML-792. While most were resistant, seven displayed intermediate sensitivity, and a further five were particularly sensitive, with sensitivity accompanied by mitotic errors, polyploidy, apoptosis, and PML body expansion. Resistance was linked to ABCB1 upregulation, and inhibiting drug efflux sensitized eight resistant OCMs. MYC target genes were enriched in sensitive models, consistent with MYC being a potential driver of response. SUMO inhibition induced an adaptive transcriptional response in resistant cells, but this was attenuated in MYC-overexpressing cells, raising the possibility that transcriptional interference disrupts the homeostatic controls required to buffer the inhibition of SUMO signaling. SUMO sensitivity did not overlap with PARP inhibitor sensitivity, supporting the therapeutic potential of apex SUMO inhibitors to target a subset of homologous-recombination-proficient ovarian cancers.
Keywords: Cancer; Molecular biology; Transcriptomics.
© 2025 The Authors.
Conflict of interest statement
The authors have no competing interests to declare that are relevant to the content of this article.
Figures







References
LinkOut - more resources
Full Text Sources
Miscellaneous
