

Chemosynthesis enhances net primary production and nutrient cycling in a hypersaline microbial mat
- PMID: 40488405
- PMCID: PMC12218205
- DOI: 10.1093/ismejo/wraf117
Chemosynthesis enhances net primary production and nutrient cycling in a hypersaline microbial mat
Abstract
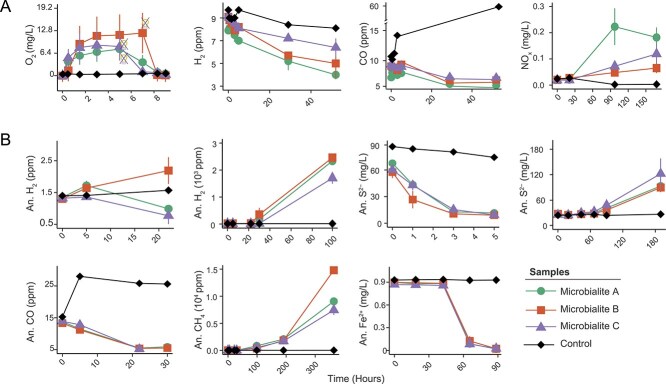
Photosynthetic microbial mats are macroscopic microbial ecosystems consisting of a wide array of functional groups and microenvironments arranged along variable redox gradients. Light energy ultimately drives primary production and a cascade of daisy-chained metabolisms. Heterotrophic members of these communities remineralise organic material, decreasing net primary production, and returning nutrients to the aqueous phase. However, reduced inorganic and one-carbon substrates such as trace gases and those released as metabolic byproducts in deeper anoxic regions of the mat, could theoretically also fuel carbon fixation, mitigating carbon loss from heterotrophy and enhancing net primary production. Here, we investigated the intricate metabolic synergies that sustain community nutrient webs in a biomineralising microbial mat from a hypersaline lake. We recovered 331 genomes spanning 40 bacterial and archaeal phyla that influence the biogeochemistry of these ecosystems. Phototrophy is a major metabolism found in 17% of the genomes, but over 50% encode enzymes to harness energy from inorganic substrates and 12% co-encode chemosynthetic carbon fixation pathways that use sulfide and hydrogen as electron donors. We experimentally demonstrated that the microbial community oxidises ferrous iron, ammonia, sulfide, and reduced trace gas substrates aerobically and anaerobically. Furthermore, carbon isotope assays revealed that diverse chemosynthetic pathways contribute significantly to carbon fixation and organic matter production alongside photosynthesis. Chemosynthesis in microbial mats results from a complex suite of spatially organised metabolic interactions and continuous nutrient cycling, which decouples carbon fixation from the diurnal cycle, and enhances the net primary production of these highly efficient ecosystems.
Keywords: carbon fixation; chemosynthesis; extreme environments; isotopes; metabolic interactions; metagenomics; microbial mat; microbialites.
© The Author(s) 2025. Published by Oxford University Press on behalf of the International Society for Microbial Ecology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Walter MR, Buick R, Dunlop JSR. Stromatolites 3,400–3,500 Myr old from the north pole area. Western Australia Nature 1980;284:443–5.
-
- Van Kranendonk MJ, Philippot P, Lepot K. et al. Geological setting of Earth’s oldest fossils in the ca. 3.5 Ga dresser formation, Pilbara craton, Western Australia. Precambrian Res 2008;167:93–124.
-
- Dick GJ, Grim SL, Klatt JM. Controls on O 2 production in cyanobacterial Mats and Implications for Earth’s oxygenation. Annu Rev Earth Planet Sci 2018;46:123–47.
-
- Nisbet EG, Fowler CMR. Archaean metabolic evolution of microbial mats. Proc R Soc Lond B 1999;266:2375–82.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
