

Enhancing titers of therapeutic lentiviral vectors using PKC agonists
- PMID: 40491664
- PMCID: PMC12146000
- DOI: 10.1016/j.omtm.2025.101484
Enhancing titers of therapeutic lentiviral vectors using PKC agonists
Abstract
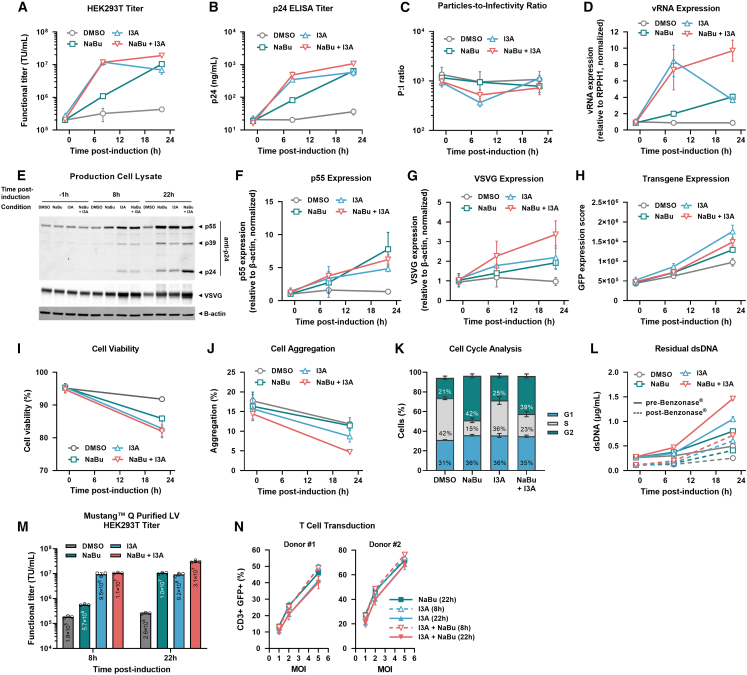
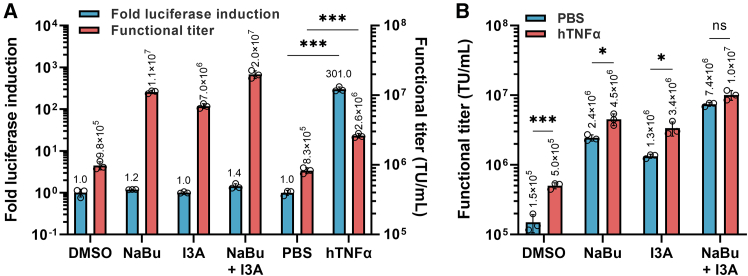
Lentiviral vector (LV)-based therapies employ the molecular machinery of HIV-1 to stably integrate therapeutic genes into patient cells for long-term disease correction. However, suboptimal expression of LV components in HEK293T-based production systems can limit titers and hinder clinical product development. Here, we identify protein kinase C (PKC) agonists as robust enhancers of LV production. PKC activation resulted in rapid transcription of LV genomic RNA and accelerated vector particle release in a manner that complemented the use of the histone deacetylase (HDAC) inhibitor, sodium butyrate. Stimulation of HEK293T cells strongly upregulated AP-1 transcription factor subunits independently of nuclear factor κB (NF-κB) pathway activation. Application of PKC agonists in LV production resulted in a ∼3-fold improvement in the titer of a chimeric antigen receptor (CAR)-LV. Furthermore, a ∼9-fold increase in titer was achieved when this induction method was combined with co-expression of an LV RNA-targeted U1 snRNA enhancer. Importantly, LV produced using PKC agonists had comparable particle-to-infectivity ratios and preserved T cell transduction efficiency. These findings suggest that incorporating PKC agonists into commercial LV manufacturing could considerably reduce the cost per patient dose of new LV-based gene therapies.
Keywords: LV manufacturing; PKC; PKC agonist; U1 snRNA-based lentiviral vector enhancer; bioprocessing; chimeric antigen receptor T cell therapy; gene therapy; lentiviral vector; protein kinase C.
© 2025 The Author(s).
Conflict of interest statement
The work described was fully funded by Oxford Biomedica (UK) Ltd., and all authors at the time of submission were employees and hold stock or stock options within the company. The authors declare that a patent application has been submitted related to this work (patent application no. WO/2021/229242), assigned to Oxford Biomedica (UK) Ltd.
Figures





References
-
- de Vries W., Haasnoot J., van der Velden J., van Montfort T., Zorgdrager F., Paxton W., Cornelissen M., van Kuppeveld F., de Haan P., Berkhout B. Increased virus replication in mammalian cells by blocking intracellular innate defense responses. Gene Ther. 2008;15:545–552. doi: 10.1038/gt.2008.12. - DOI - PubMed
LinkOut - more resources
Full Text Sources
