

This is a preprint.
Optimization and variability can coexist
- PMID: 40492252
- PMCID: PMC12148085
Optimization and variability can coexist
Abstract
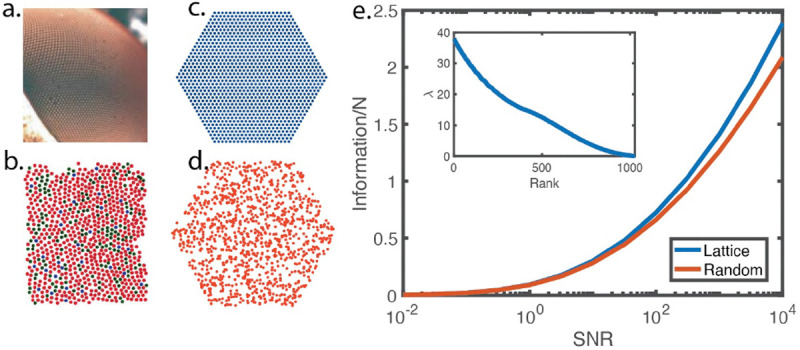
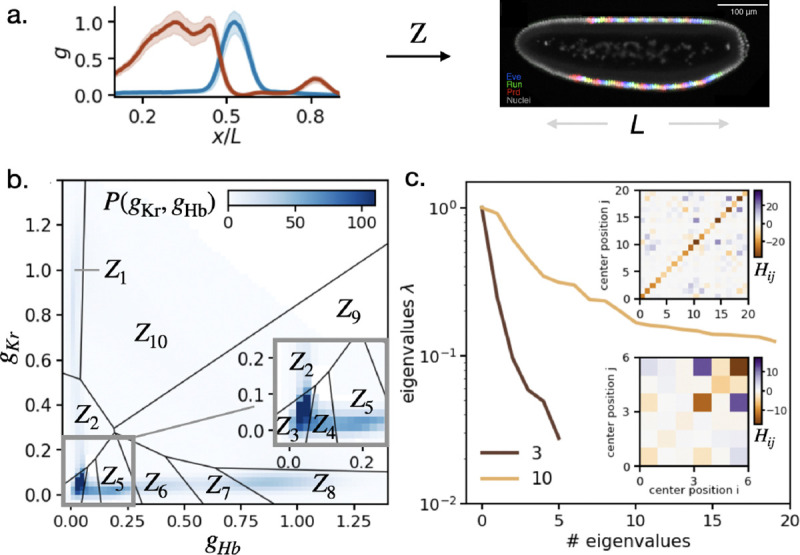
Many biological systems perform close to their physical limits, but promoting this optimality to a general principle seems to require implausibly fine tuning of parameters. Using examples from a wide range of systems, we show that this intuition is wrong. Near an optimum, functional performance depends on parameters in a "sloppy" way, with some combinations of parameters being only weakly constrained. Absent any other constraints, this predicts that we should observe widely varying parameters, and we make this precise: the entropy in parameter space can be extensive even if performance on average is very close to optimal. This removes a major objection to optimization as a general principle, and rationalizes the observed variability.
Figures



References
-
- Rieke F. & Baylor D. A. Single photon detection by rod cells of the retina. Reviews of Modern Physics 70, 1027–1036 (1998).
-
- Barlow H. B. The size of ommatidia in apposition eyes. Journal of Experimental Biology 29, 667–674 (1952).
-
- Snyder A. W. Acuity of compound eyes: Physical limitations and design. Journal of Comparative Physiology 116, 161–182 (1977).
-
- Bialek W. Biophysics: Searching for Principles (Princeton University Press, Princeton NJ, 2012).
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources