

Identification of the AKCDPK gene family and AkCDPK15 functional analysis under drought and salt stress
- PMID: 40498756
- PMCID: PMC12157117
- DOI: 10.1371/journal.pone.0325453
Identification of the AKCDPK gene family and AkCDPK15 functional analysis under drought and salt stress
Abstract
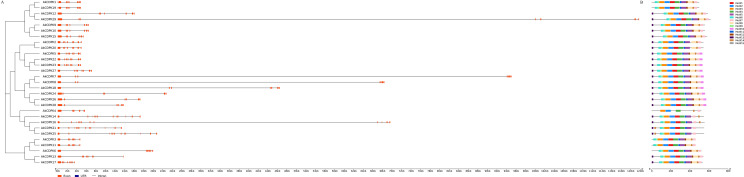
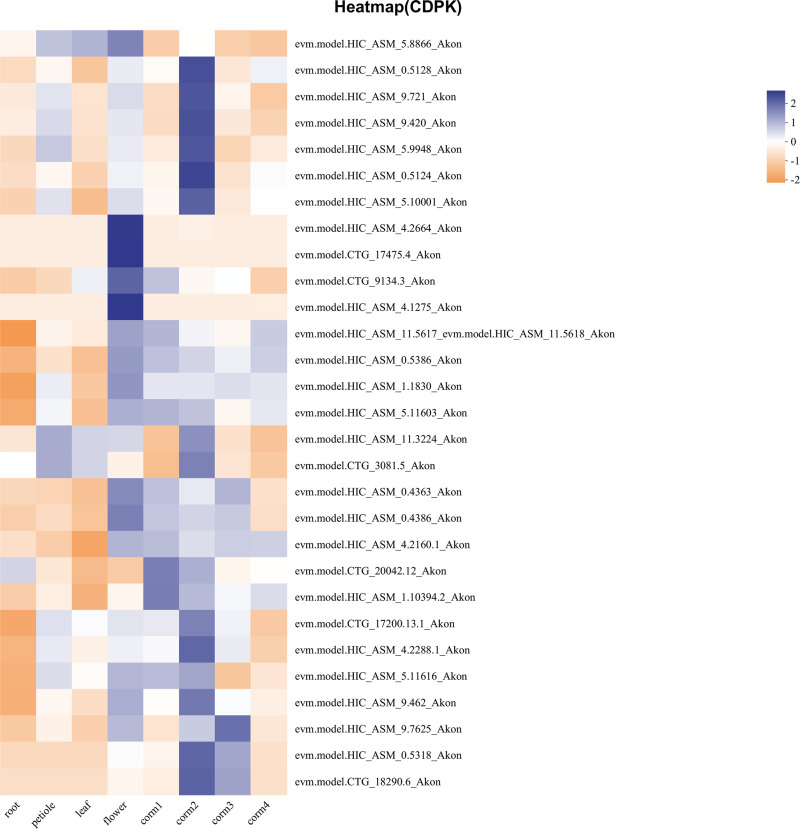
Konjac is one of the important economic crops for poverty alleviation in mountainous areas of Yunnan Province, China. However, there are always various biotic and abiotic stress during its growth, leading to production reduction and quality decline. Calcium-dependent protein kinases (CDPKs) are an important class of genes involved in calcium ion signal transmission within plant tissue cells, yet their presence and functions in konjac remain unexplored. This study aimed to identify the members of the AkCDPK gene family in the Amorphophallus konjac genome and understand their evolution and responses to various stresses. A total of 29 AkCDPK genes were identified and categorized into four subgroups that unevenly distributed across 12 chromosomes. Most AkCDPK have undergone purifying selection during evolution. Cis-acting element analysis revealed that several AkCDPK are involved in phytohormone induction, defence, stress response, and plant development. Expression analysis indicated tissue specificity, and responses to salt, drought, and Pectobacterium carotovorum subsp. carotovorum stress. AkCDPK15, encoding 582 amino acids, was cloned. AkCDPK15 was mainly localised on the cell membrane, and overexpression in tobacco revealed that it can positively regulate the tolerance of transgenic tobacco strains to salt and drought stress. These findings provide a theoretical foundation for future research on the function of the CDPK gene family in A. konjac, potentially aiding in the development of stress-resistant konjac varieties.
Copyright: © 2025 Gao et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures









Similar articles
-
Genome-wide identification and characterization of Calcium-Dependent Protein Kinase (CDPK) gene family in autotetraploid cultivated alfalfa (Medicago sativa subsp. sativa) and expression analysis under abiotic stresses.BMC Plant Biol. 2024 Dec 23;24(1):1241. doi: 10.1186/s12870-024-05993-7. BMC Plant Biol. 2024. PMID: 39716096 Free PMC article.
-
Genome-wide identification and evolution of the SAP gene family in sunflower (Helianthus annuus L.) and expression analysis under salt and drought stress.PeerJ. 2024 Jul 30;12:e17808. doi: 10.7717/peerj.17808. eCollection 2024. PeerJ. 2024. PMID: 39099650 Free PMC article.
-
Genome-wide analysis of the Amorphophallus konjac AkCSLA gene family and its functional characterization in drought tolerance of transgenic arabidopsis.BMC Plant Biol. 2024 Oct 31;24(1):1033. doi: 10.1186/s12870-024-05747-5. BMC Plant Biol. 2024. PMID: 39478464 Free PMC article.
-
[Identification and expression analysis of the HSP70 gene family under abiotic stresses in Litchi chinensis].Sheng Wu Gong Cheng Xue Bao. 2024 Apr 25;40(4):1102-1119. doi: 10.13345/j.cjb.230450. Sheng Wu Gong Cheng Xue Bao. 2024. PMID: 38658152 Chinese.
-
[Functions and mechanisms of CDPKs in plant responses to abiotic stress].Sheng Wu Gong Cheng Xue Bao. 2024 Oct 25;40(10):3337-3359. doi: 10.13345/j.cjb.230827. Sheng Wu Gong Cheng Xue Bao. 2024. PMID: 39467737 Review. Chinese.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
