

Cell states and neighborhoods in distinct clinical stages of primary and metastatic esophageal adenocarcinoma
- PMID: 40499545
- PMCID: PMC12208317
- DOI: 10.1016/j.xcrm.2025.102188
Cell states and neighborhoods in distinct clinical stages of primary and metastatic esophageal adenocarcinoma
Abstract
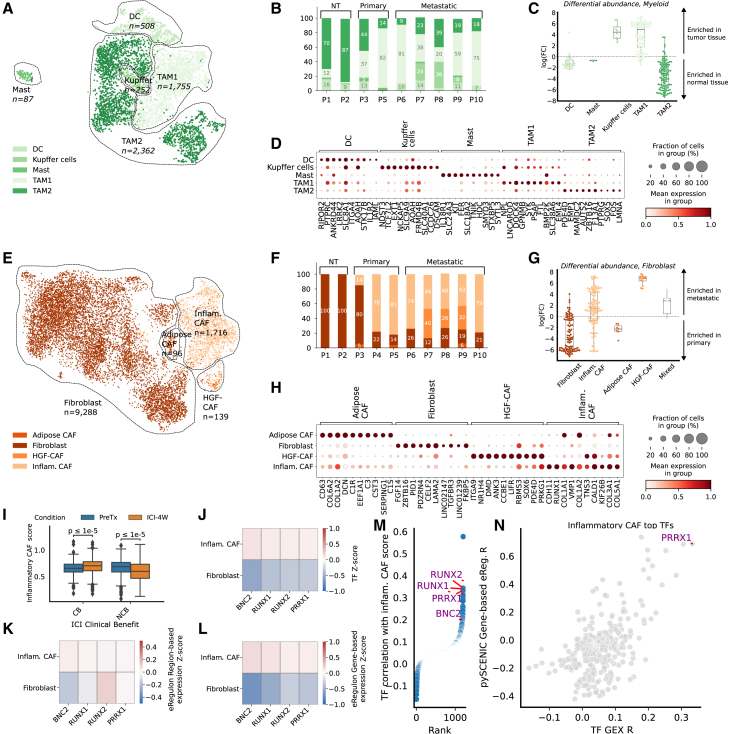
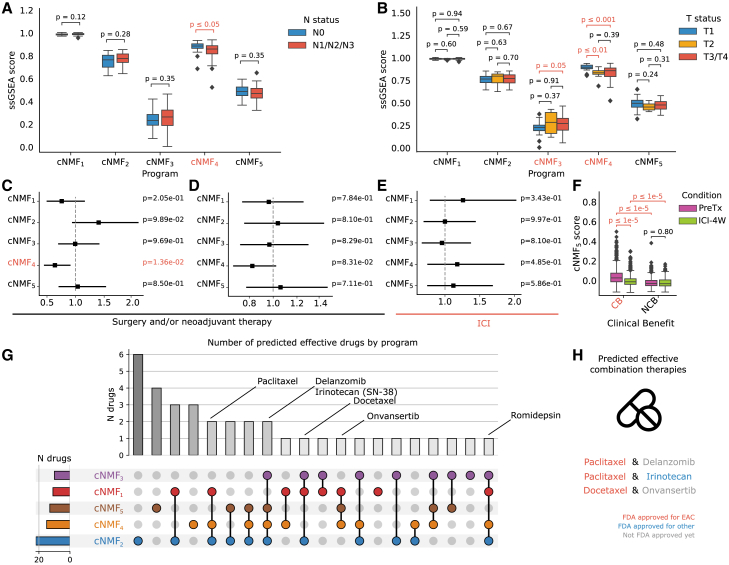
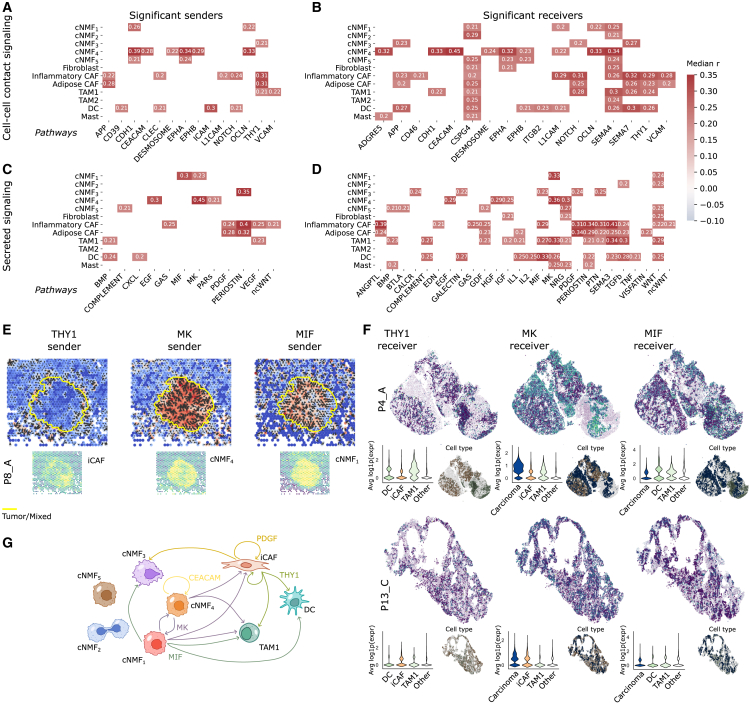
Esophageal adenocarcinoma (EAC) is a highly lethal cancer of the upper gastrointestinal tract with rising incidence in western populations. To decipher EAC disease progression and therapeutic response, we perform multiomic analyses of a cohort of primary and metastatic EAC tumors, incorporating single-nuclei transcriptomic and chromatin accessibility sequencing along with spatial profiling. We recover tumor microenvironmental features previously described to associate with therapy response. We subsequently identify five malignant cell programs, including undifferentiated, intermediate, differentiated, epithelial-to-mesenchymal transition, and cycling programs, which are associated with differential epigenetic plasticity and clinical outcomes, and for which we infer candidate transcription factor regulons. Furthermore, we reveal diverse spatial localizations of malignant cells expressing their associated transcriptional programs and predict their significant interactions with microenvironmental cell types. We validate our findings in three external single-cell RNA sequencing (RNA-seq) and three bulk RNA-seq studies. Altogether, our findings advance the understanding of EAC heterogeneity, disease progression, and therapeutic response.
Keywords: bioinformatics; computational biology; epigenetics; esophageal adenocarcinoma; gastrointestinal cancer; oncology; single cell; spatial transcriptomics; transcriptomics.
Copyright © 2025 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.M.V.A.—advisory/consulting: Enara Bio, Manifold Bio, Monte Rosa, Novartis Institute for Biomedical Research, Serinus Bio, and TracerDx; research support: Novartis, BMS, Sanofi, and NextPoint; equity: Tango Therapeutics, Genome Medical, Genomic Life, Enara Bio, Manifold Bio, Microsoft, Monte Rosa, Riva Therapeutics, Serinus Bio, Syapse, and TracerDx; travel reimbursement: none; patents: institutional patents filed on chromatin mutations and immunotherapy response, and methods for clinical interpretation; intermittent legal consulting on patents for Foaley & Hoag; editorial boards: Science Advances. A.J.A. has consulted for Anji Pharmaceuticals, Affini-T Therapeutics, Arrakis Therapeutics, AstraZeneca, Boehringer Ingelheim, Kestrel Therapeutics, Merck & Co., Inc., Mirati Therapeutics, Nimbus Therapeutics, Oncorus, Inc., Plexium, Quanta Therapeutics, Revolution Medicines, Reactive Biosciences, Riva Therapeutics, Servier Pharmaceuticals, Syros Pharmaceuticals, T-knife Therapeutics, Third Rock Ventures, and Ventus Therapeutics. A.J.A. holds equity in Riva Therapeutics and Kestrel Therapeutics. A.J.A. has research funding from Amgen, AstraZeneca, Boehringer Ingelheim, Bristol Myers Squibb, Deerfield, Inc., Eli Lilly, Mirati Therapeutics, Nimbus Therapeutics, Novartis, Novo Ventures, Revolution Medicines, and Syros Pharmaceuticals.
Figures





Update of
-
Cell states and neighborhoods in distinct clinical stages of primary and metastatic esophageal adenocarcinoma.bioRxiv [Preprint]. 2025 Mar 3:2024.08.17.608386. doi: 10.1101/2024.08.17.608386. bioRxiv. 2025. Update in: Cell Rep Med. 2025 Jun 17;6(6):102188. doi: 10.1016/j.xcrm.2025.102188. PMID: 39229240 Free PMC article. Updated. Preprint.
References
-
- Nowicki-Osuch K., Zhuang L., Jammula S., Bleaney C.W., Mahbubani K.T., Devonshire G., Katz-Summercorn A., Eling N., Wilbrey-Clark A., Madissoon E., et al. Molecular phenotyping reveals the identity of Barrett’s esophagus and its malignant transition. Science. 2021;373:760–767. - PubMed
-
- Pennathur A., Gibson M.K., Jobe B.A., Luketich J.D. Oesophageal carcinoma. Lancet. 2013;381:400–412. - PubMed
-
- Derakhshan M.H., Arnold M., Brewster D.H., Going J.J., Mitchell D.R., Forman D., McColl K.E.L. Worldwide Inverse Association between Gastric Cancer and Esophageal Adenocarcinoma Suggesting a Common Environmental Factor Exerting Opposing Effects. Am. J. Gastroenterol. 2016;111:228–239. - PubMed
MeSH terms
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
