

Developmental trajectory and evolutionary origin of thymic mimetic cells
- PMID: 40500437
- PMCID: PMC12286861
- DOI: 10.1038/s41586-025-09148-y
Developmental trajectory and evolutionary origin of thymic mimetic cells
Abstract
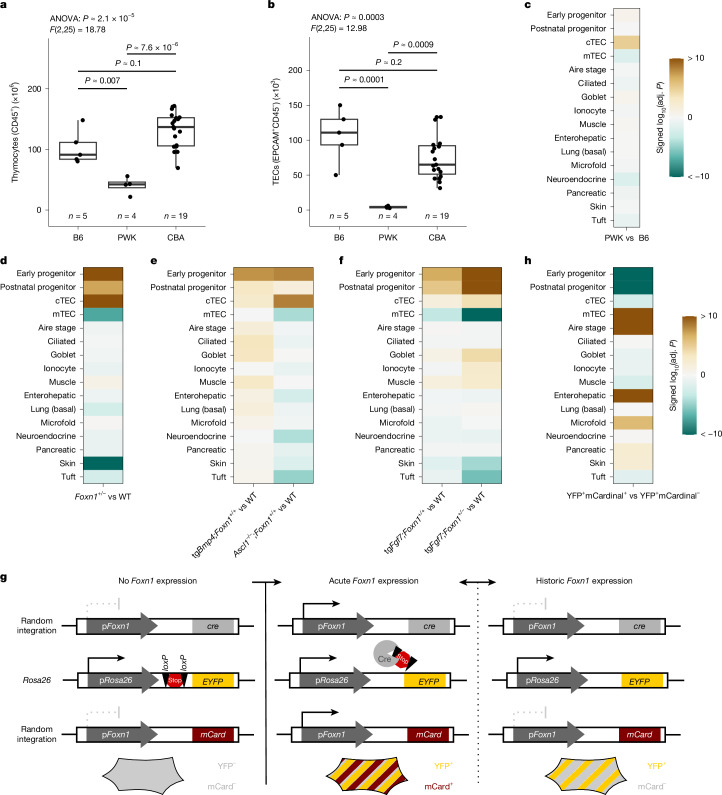
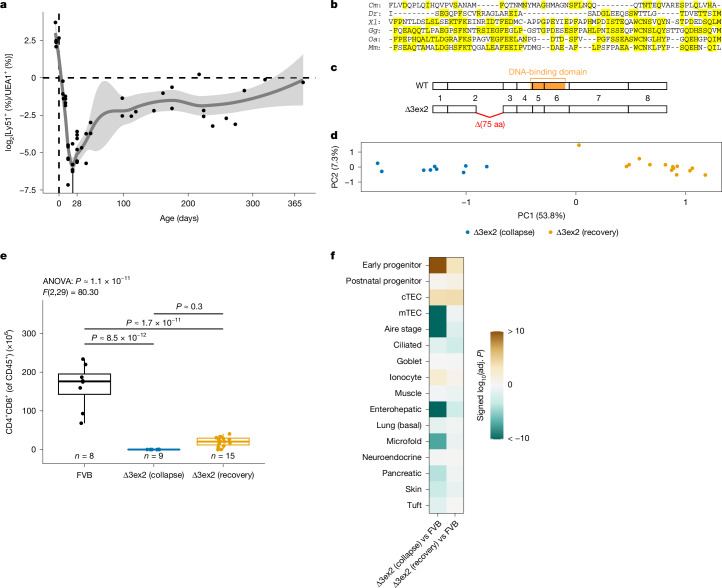
The generation of self-tolerant repertoires of T cells depends on the expression of peripheral self antigens in the thymic epithelium1 and the presence of small populations of cells that mimic the diverse phenotypes of peripheral tissues2-7. Whereas the molecular underpinnings of self-antigen expression have been extensively studied8, the developmental origins and differentiation pathways of thymic mimetic cells remain to be identified. Moreover, the histological identification of myoid and other peripheral cell types as components of the thymic microenvironment of many vertebrate species9 raises questions regarding the evolutionary origin of this unique tolerance mechanism. Here we show that during mouse development, mimetic cells appear in the microenvironment in two successive waves. Cells that exhibit transcriptional signatures characteristic of muscle, ionocyte, goblet and ciliated cells emerge before birth, whereas others, such as those that mimic enterohepatic cells and skin keratinocytes, appear postnatally. These two groups also respond differently to modulations of thymic epithelial cell progenitor pools caused by deletions of Foxn1 and Ascl1, expression of a hypomorphic variant of the transcription factor FOXN1, and overexpression of the signalling molecules BMP4 and FGF7. Differences in mimetic cell populations were also observed in thymic microenvironments reconstructed by replacement of mouse Foxn1 with evolutionarily ancient Foxn1/4 gene family members, including the Foxn4 gene of the cephalochordate amphioxus and the Foxn4 and Foxn1 genes of a cartilaginous fish. Whereas some cell types, such as ciliated cells, develop in the thymus in the absence of FOXN1, mimetic cells that appear postnatally, such as enterohepatic cells, require the activity of the vertebrate-specific transcription factor FOXN1. The thymus of cartilaginous fishes and the thymoid of lampreys, a representative of jawless vertebrates, which exhibit an alternative adaptive immune system10, also harbour cells that express genes encoding peripheral tissue components such as the liver-specific protein transthyretin. Our findings suggest an evolutionary model of successive changes of thymic epithelial genetic networks enabling the coordinated contribution of peripheral antigen expression and mimetic cell formation to achieve central tolerance for vertebrate-specific innovations of tissues such as the liver11,12.
© 2025. The Author(s).
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures














References
-
- Derbinski, J., Schulte, A., Kyewski, B. & Klein, L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol.2, 1032–1039 (2001). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
