

This is a preprint.
Hormetic elevation of taurine restrains inflammaging by deactivating the NLRP3 inflammasome
- PMID: 40501605
- PMCID: PMC12154967
- DOI: 10.1101/2025.05.27.656381
Hormetic elevation of taurine restrains inflammaging by deactivating the NLRP3 inflammasome
Abstract
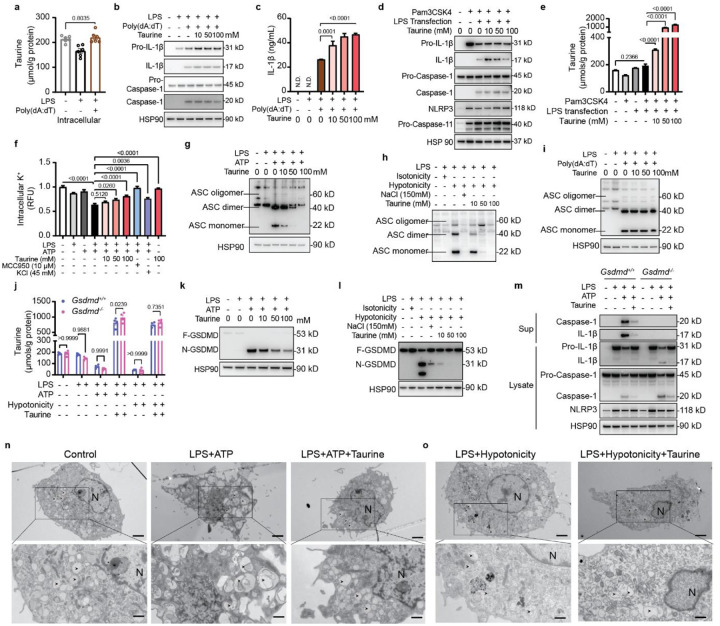
Taurine, the most abundant sulfonic amino acid in humans is largely obtained from diets rich in animal proteins. However, taurine is dietary non-essential because it can be synthesized from cysteine by activation of transsulfuration pathway (TSP) when food consumption is low or if the diet is predominantly plant based. The decline of taurine was proposed as the driver of aging through an undefined mechanism. Here, we found that mild food restriction in humans for one year that resulted in 14% reduction of calorie intake elevated the hypotaurine and taurine concentration in adipose tissue. Therefore, we investigated whether elevated taurine mimics caloric-restriction's beneficial effects on inflammation, a key mechanism of aging. Interestingly, aging increased the circulating and tissue concentrations of taurine suggesting that elevated taurine may serve as a hormetic stress response metabolite that regulates mechanism of age-related inflammation. The elevated taurine protected mice against mortality from sepsis and inhibited inflammasome-driven inflammation and gasdermin-D (GSDMD) mediated pyroptosis. Mechanistically, 'danger signals' including hypotonicity that activate NLRP3-inflammasome, caused upstream taurine efflux from macrophages, which triggered potassium (K+) release and downstream canonical NLRP3 inflammasome assembly, caspase-1 activation, GSDMD cleavage and IL-1β and IL-18 secretion that was reversed by taurine restoration. Notably, taurine does not efflux from GSDMD pore and inhibited IL-1β from macrophages independently of known transporters SLC6A6 and SLC36A1. Increased taurine in old mice promotes healthspan by inducing anti-inflammatory pathways previously linked to youthfulness. These findings demonstrate that taurine is an upstream metabolic sensor of cellular perturbations that control NLRP3 inflammasome and lowers age-related inflammation.
Figures




References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous