

This is a preprint.
Geometrically encoded positioning of introns, intergenic segments, and exons in the human genome
- PMID: 40501616
- PMCID: PMC12154594
- DOI: 10.1101/2025.05.29.656862
Geometrically encoded positioning of introns, intergenic segments, and exons in the human genome
Abstract
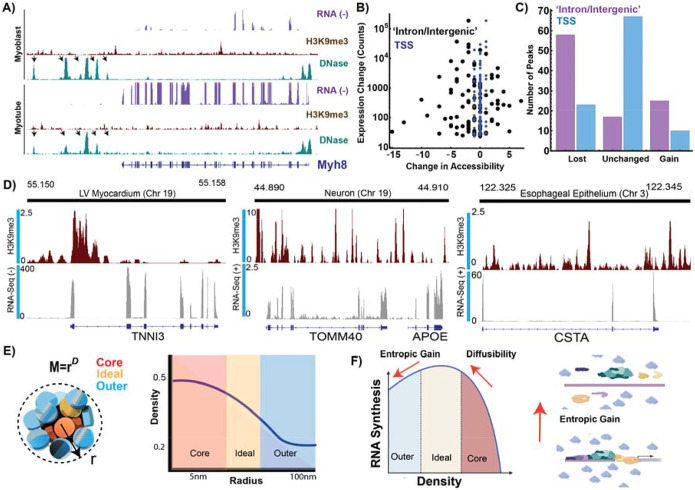
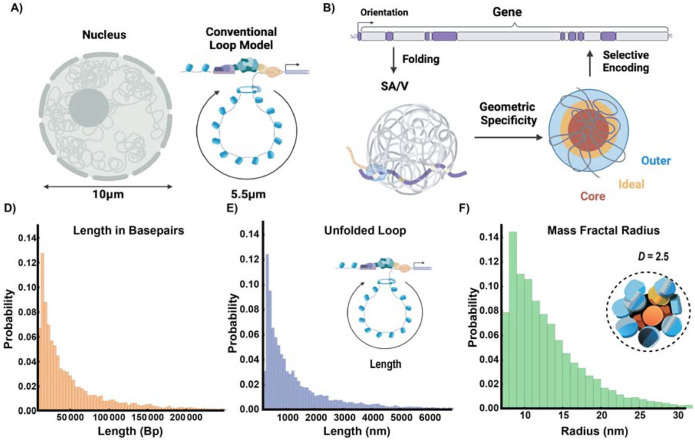
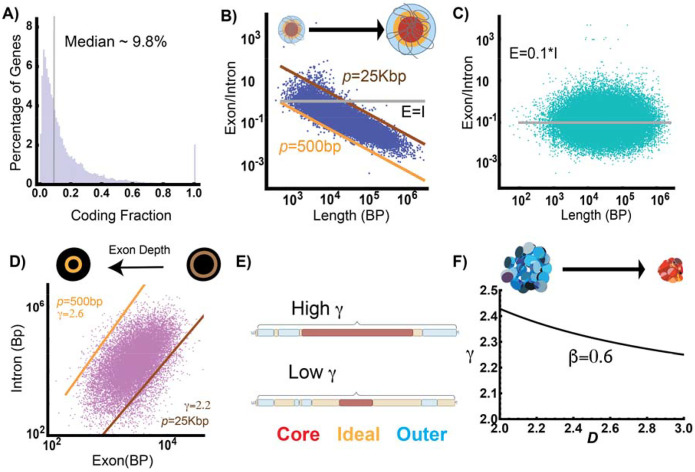
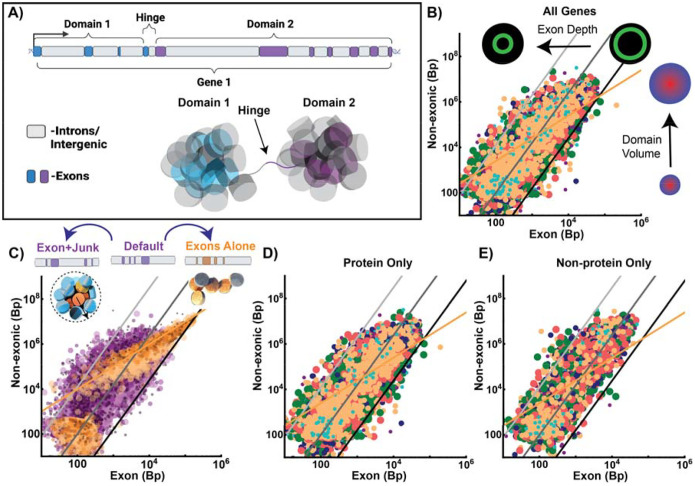
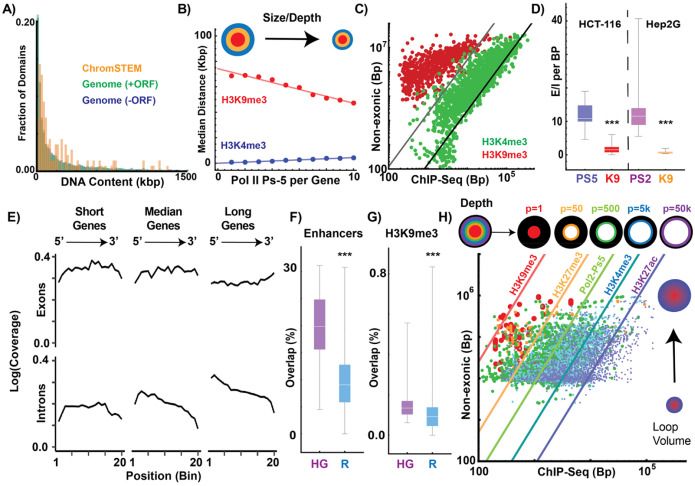
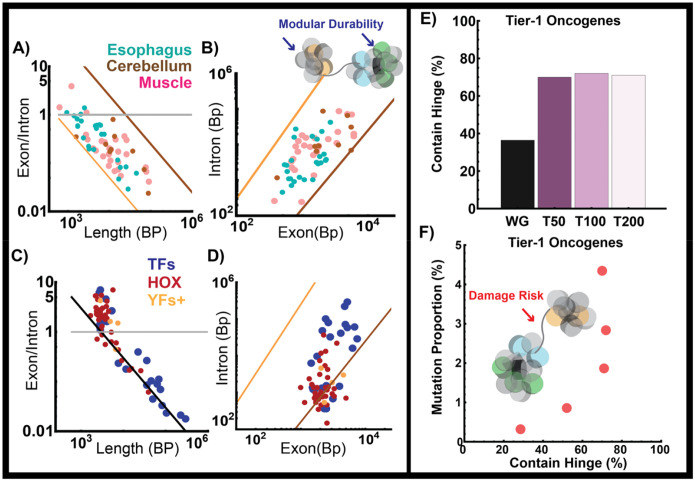
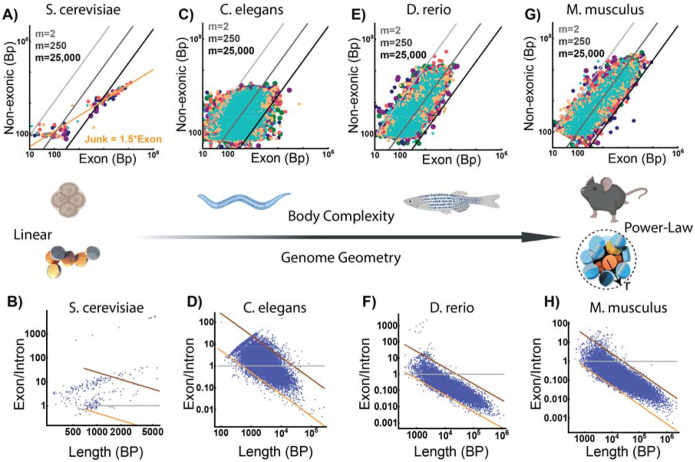
Human tissues require a mechanism to generate durable, yet modifiable, transcriptional memories to sustain cell function across a lifetime. Previously, we demonstrated that nanoscale packing domains couple heterochromatin (cores) and euchromatin (outer zone) into unified reaction volumes that can generate transcriptional memory. In prior work, this framework demonstrated that RNA synthesis occurred within the ideal zone (intermediate density) portions of the domain. Naturally, this creates a question of where genes are positioned in relation to the packing domain architecture and which genetic material fills the domain core to sustain transcription. Here we propose that this could be solved by the encoded positioning of introns, intergenic segments, and exons as a projection of the functional packing layers of domains. This suggests that introns and intergenic segments are coupled to adjacent exons to generate coherent packing domain volumes. We illustrate how this organization would reconcile contradictions in epigenetic patterns, non-randomness in oncogenic mutations, and produce durable transcriptional memory. We conclude by showing that this genome geometry might have coincided with the rapid evolution of body-plan complexity, suggesting that chromatin geometry could be fundamental to metazoan evolution.
Conflict of interest statement
Competing Interest Statement: The authors declare no financial interests or consulting interests related to this work.
Figures
Similar articles
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
Adapting Safety Plans for Autistic Adults with Involvement from the Autism Community.Autism Adulthood. 2025 May 28;7(3):293-302. doi: 10.1089/aut.2023.0124. eCollection 2025 Jun. Autism Adulthood. 2025. PMID: 40539213
-
Can a Liquid Biopsy Detect Circulating Tumor DNA With Low-passage Whole-genome Sequencing in Patients With a Sarcoma? A Pilot Evaluation.Clin Orthop Relat Res. 2025 Jan 1;483(1):39-48. doi: 10.1097/CORR.0000000000003161. Epub 2024 Jun 21. Clin Orthop Relat Res. 2025. PMID: 38905450
-
Factors that influence parents' and informal caregivers' views and practices regarding routine childhood vaccination: a qualitative evidence synthesis.Cochrane Database Syst Rev. 2021 Oct 27;10(10):CD013265. doi: 10.1002/14651858.CD013265.pub2. Cochrane Database Syst Rev. 2021. PMID: 34706066 Free PMC article.
-
Developing a role for patients and the public in the implementation of health and social care research evidence into practice: the PIPER study (Pathways to Implementation for Public Engagement in Research) realist evaluation protocol.Res Involv Engagem. 2025 Jul 14;11(1):80. doi: 10.1186/s40900-025-00728-w. Res Involv Engagem. 2025. PMID: 40660365 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources