

This is a preprint.
Identification of chromatin-associated RNAs at human centromeres
- PMID: 40501620
- PMCID: PMC12157442
- DOI: 10.1101/2025.06.05.658139
Identification of chromatin-associated RNAs at human centromeres
Abstract
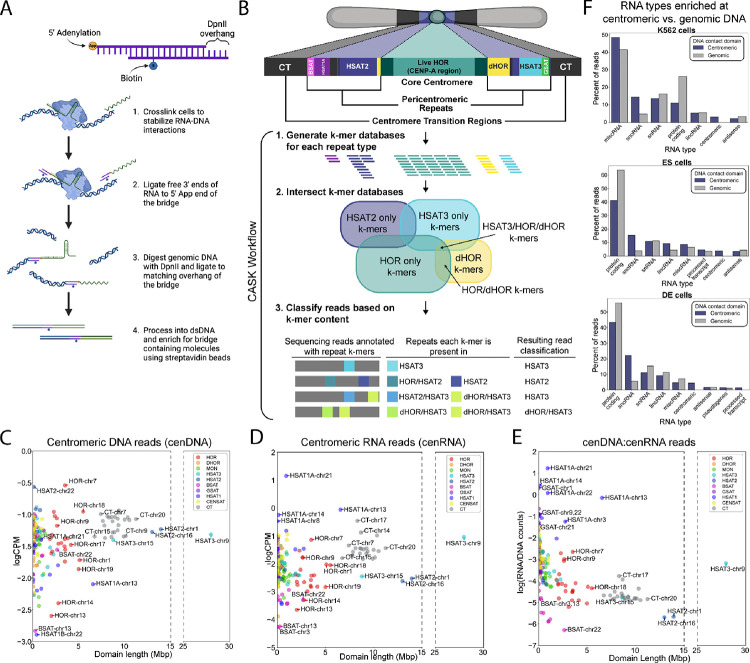
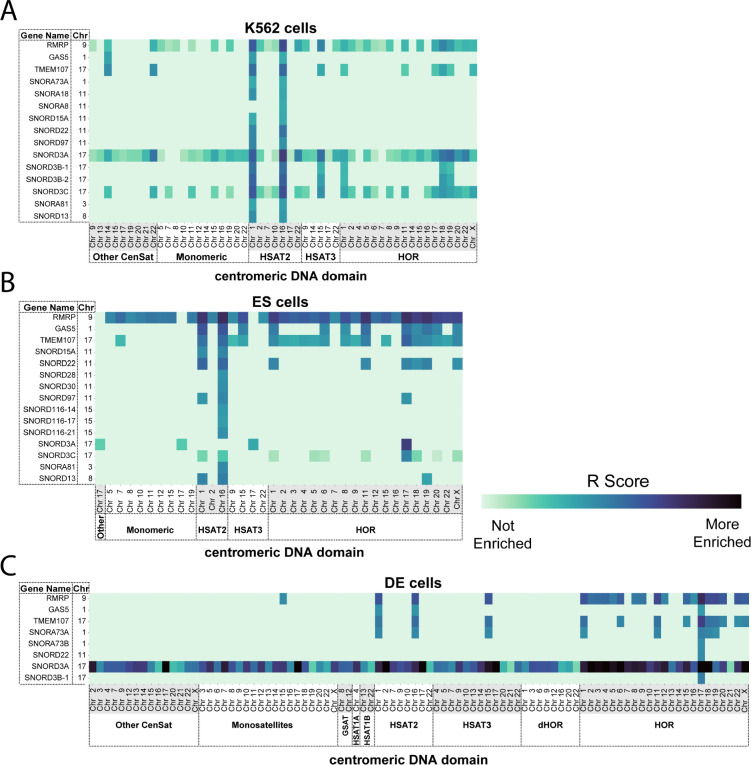
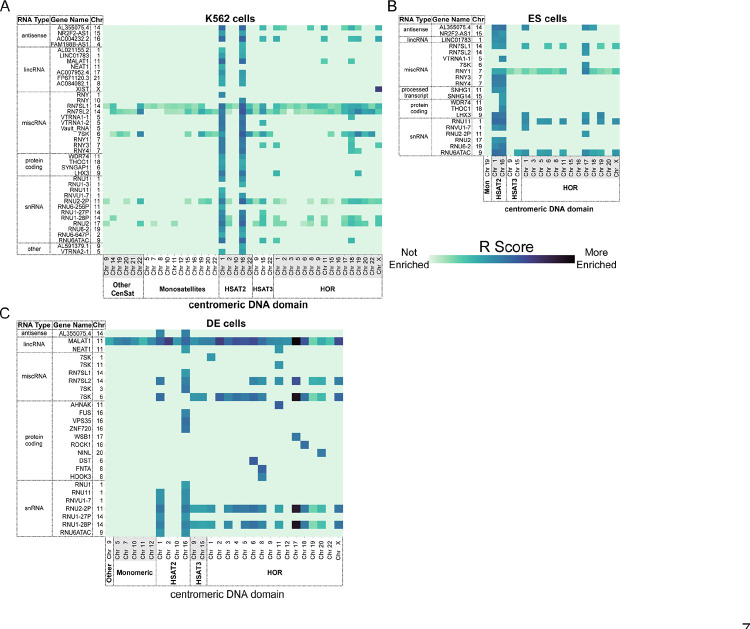
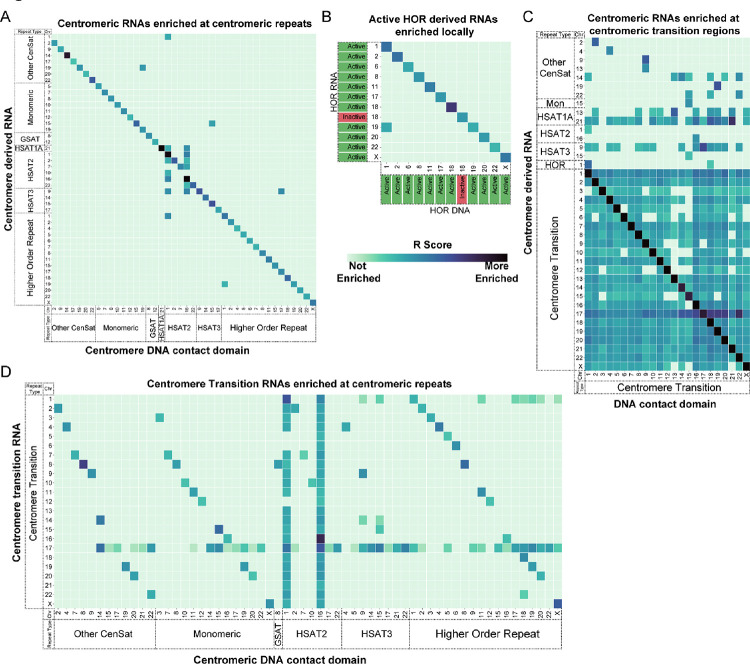
Centromeres are a specialized chromatin domain that are required for the assembly of the mitotic kinetochore and the accurate segregation of chromosomes. Non-coding RNAs play essential roles in regulating genome organization including at the unique chromatin environment present at human centromeres. We performed Chromatin-Associated RNA sequencing (ChAR-seq) in three different human cell lines to identify and map RNAs associated with centromeric chromatin. Centromere enriched RNAs display distinct contact behaviors across repeat arrays and generally belong to three categories: centromere encoded, nucleolar localized, and highly abundant, broad-binding RNAs. Most centromere encoded RNAs remain locally associated with their transcription locus with the exception of a subset of human satellite RNAs. This work provides a comprehensive identification of centromere bound RNAs that may regulate the organization and activity of the centromere.
Conflict of interest statement
Declarations of interest The authors declare no competing interests.
Figures
Similar articles
-
Conservation of dichromatin organization along regional centromeres.Cell Genom. 2025 Apr 9;5(4):100819. doi: 10.1016/j.xgen.2025.100819. Epub 2025 Mar 26. Cell Genom. 2025. PMID: 40147439 Free PMC article.
-
Unveiling the distinctive traits of functional rye centromeres: minisatellites, retrotransposons, and R-loop formation.Sci China Life Sci. 2024 Sep;67(9):1989-2002. doi: 10.1007/s11427-023-2524-0. Epub 2024 May 24. Sci China Life Sci. 2024. PMID: 38805064
-
Drugs for preventing postoperative nausea and vomiting in adults after general anaesthesia: a network meta-analysis.Cochrane Database Syst Rev. 2020 Oct 19;10(10):CD012859. doi: 10.1002/14651858.CD012859.pub2. Cochrane Database Syst Rev. 2020. PMID: 33075160 Free PMC article.
-
Behavioral interventions to reduce risk for sexual transmission of HIV among men who have sex with men.Cochrane Database Syst Rev. 2008 Jul 16;(3):CD001230. doi: 10.1002/14651858.CD001230.pub2. Cochrane Database Syst Rev. 2008. PMID: 18646068
-
The effects of Non3 mutations on chromatin organization in Drosophila melanogaster.Vavilovskii Zhurnal Genet Selektsii. 2025 Jun;29(3):401-413. doi: 10.18699/vjgb-25-43. Vavilovskii Zhurnal Genet Selektsii. 2025. PMID: 40567590 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources