

This is a preprint.
Eph Receptor Tyrosine Kinases Are Functional Entry Receptors for Murine Gammaherpesvirus 68
- PMID: 40501621
- PMCID: PMC12157678
- DOI: 10.1101/2025.06.05.657996
Eph Receptor Tyrosine Kinases Are Functional Entry Receptors for Murine Gammaherpesvirus 68
Abstract
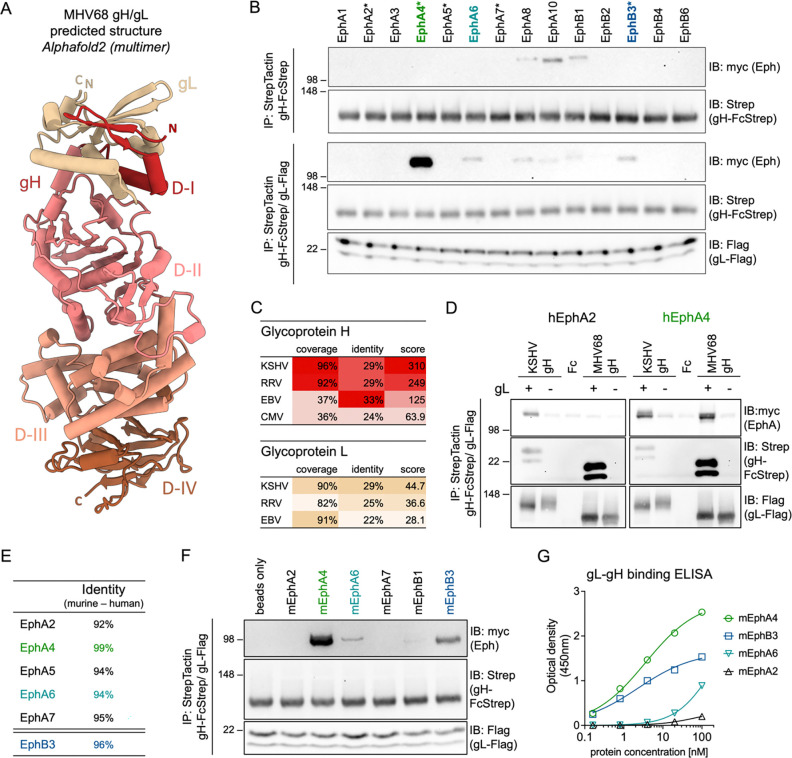
Interactions between viral glycoproteins and cellular receptors determine virus tropism and represent promising targets for vaccines. Eph receptor tyrosine kinases are conserved receptors for the human oncogenic gammaherpesviruses, Kaposi sarcoma herpesvirus (KSHV) and Epstein-Barr virus (EBV), and mediate entry into target cells by interaction with the viral gH/gL glycoprotein complex. To evaluate the use of murine gammaherpesvirus 68 (MHV68), a natural pathogen of rodents, as an in vivo model system for early events in gammaherpesvirus infection, we characterized the interaction of the MHV68 gH/gL complex with Eph receptors. We demonstrate a direct interaction of MHV68 gH/gL with EphA4 and EphB3, that is conserved between human and murine receptors. Pre-incubation of MHV68 inocula with soluble decoy receptors decreased infection of permissive fibroblasts. Ectopic expression of EphA4 and EphB3 enabled MHV68 to infect otherwise non-permissive human B cells, demonstrating EphA4 and EphB3 receptor function. Targeted mutations informed by protein structure predictions demonstrate that the MHV68 gH/gL-Eph interaction is determined by domain I (D-I) and follows structural motifs previously described in the KSHV gH/gL-EphA2 complex. The importance of gH D-I is further highlighted by the analysis of gH-targeting neutralizing antibodies. Antibody adsorption via the full gH ectodomain or gH D-I led to comparable reductions in neutralization capacity of serum from WT infected mice, indicating the Eph-binding domain is a major target for gH/gL-directed neutralizing antibodies. Our study characterizes Eph receptors as novel interaction partners and entry receptors for MHV68. Conservation of entry mechanisms provides the basis for future in vivo analyses of the contribution of Eph receptors to cell-type dependent MHV68 infection, as well as targeted strategies to prevent transmission and diseases associated with chronic infection.
Figures




Similar articles
-
Generation of chimeric forms of rhesus macaque rhadinovirus expressing KSHV envelope glycoproteins gH and gL for utilization in an NHP model of infection.J Virol. 2025 Feb 25;99(2):e0192324. doi: 10.1128/jvi.01923-24. Epub 2025 Jan 21. J Virol. 2025. PMID: 39835812 Free PMC article.
-
BALB/c mice immunized with a combination of virus-like particles incorporating Kaposi sarcoma-associated herpesvirus (KSHV) envelope glycoproteins gpK8.1, gB, and gH/gL induced comparable serum neutralizing antibody activity to UV-inactivated KSHV.Oncotarget. 2017 May 23;8(21):34481-34497. doi: 10.18632/oncotarget.15605. Oncotarget. 2017. PMID: 28404899 Free PMC article.
-
Gammaherpesvirus infection triggers the formation of tRNA fragments from premature tRNAs.mBio. 2025 Jul 9;16(7):e0087525. doi: 10.1128/mbio.00875-25. Epub 2025 May 30. mBio. 2025. PMID: 40444975 Free PMC article.
-
Growth hormone for in vitro fertilisation (IVF).Cochrane Database Syst Rev. 2021 Nov 22;11(11):CD000099. doi: 10.1002/14651858.CD000099.pub4. Cochrane Database Syst Rev. 2021. PMID: 34808697 Free PMC article.
-
EORTC guidelines for the use of erythropoietic proteins in anaemic patients with cancer: 2006 update.Eur J Cancer. 2007 Jan;43(2):258-70. doi: 10.1016/j.ejca.2006.10.014. Epub 2006 Dec 19. Eur J Cancer. 2007. PMID: 17182241
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous