

This is a preprint.
Disease-causing mutations in the G protein β5 β-propeller disrupt its chaperonin-mediated folding trajectory
- PMID: 40501749
- PMCID: PMC12154622
- DOI: 10.1101/2025.05.28.656654
Disease-causing mutations in the G protein β5 β-propeller disrupt its chaperonin-mediated folding trajectory
Abstract
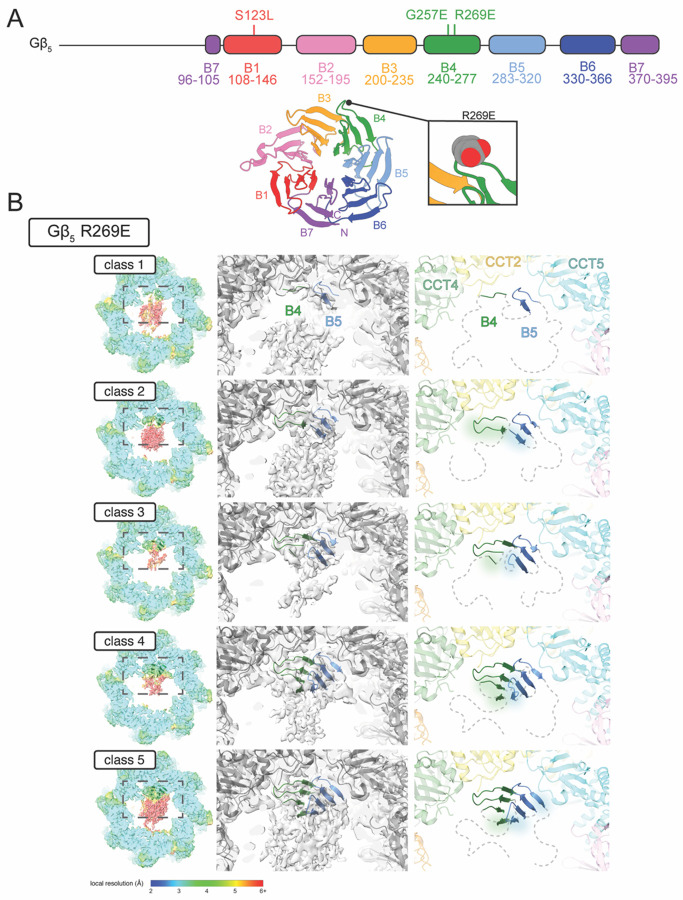
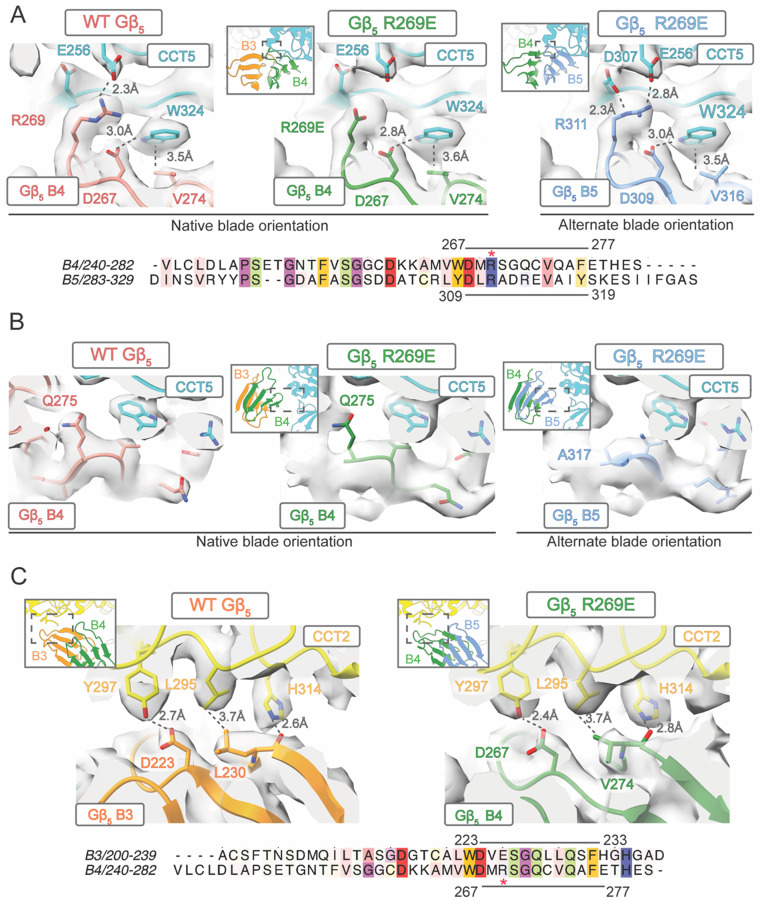
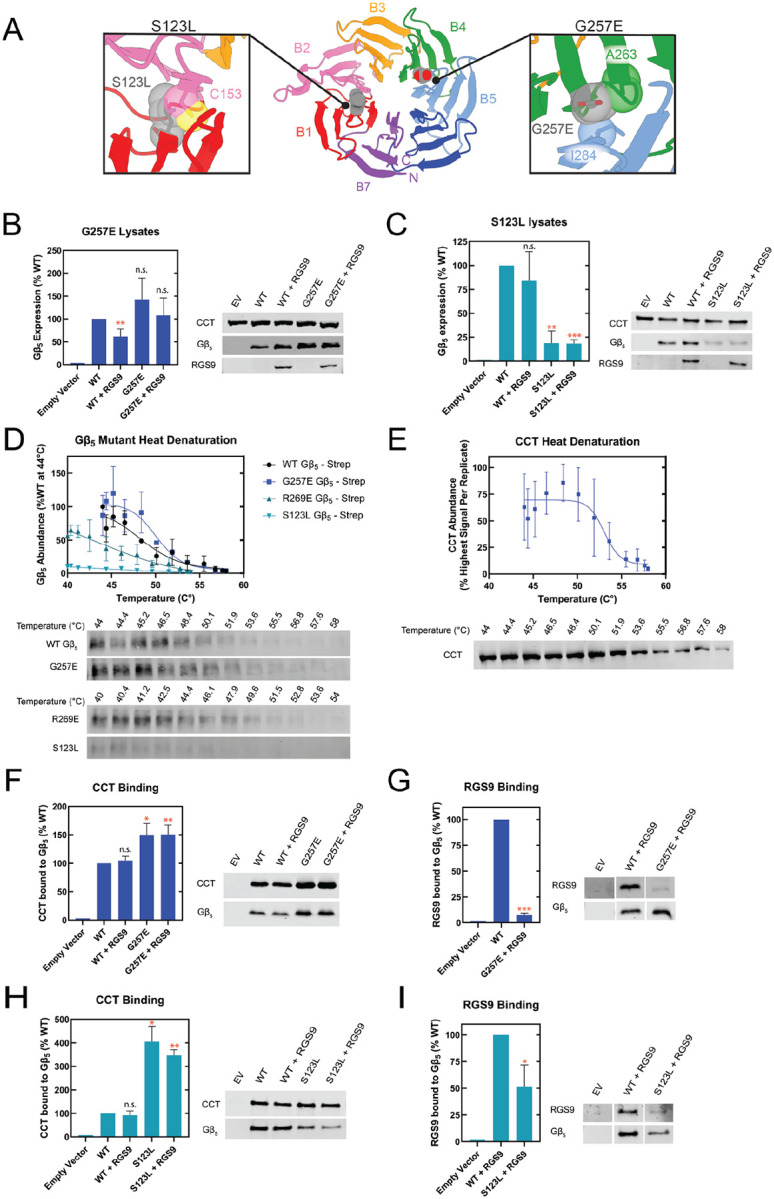
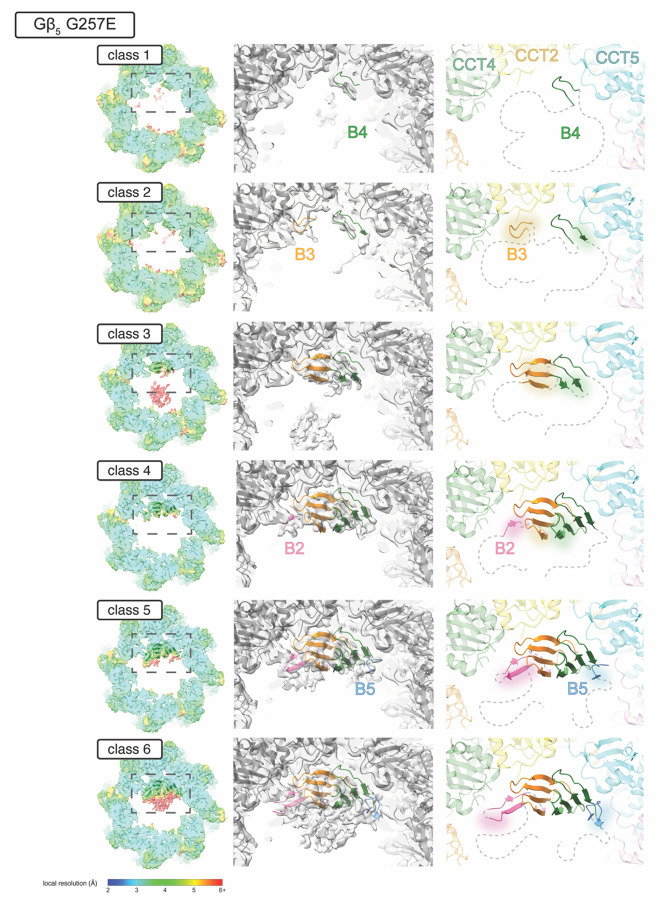
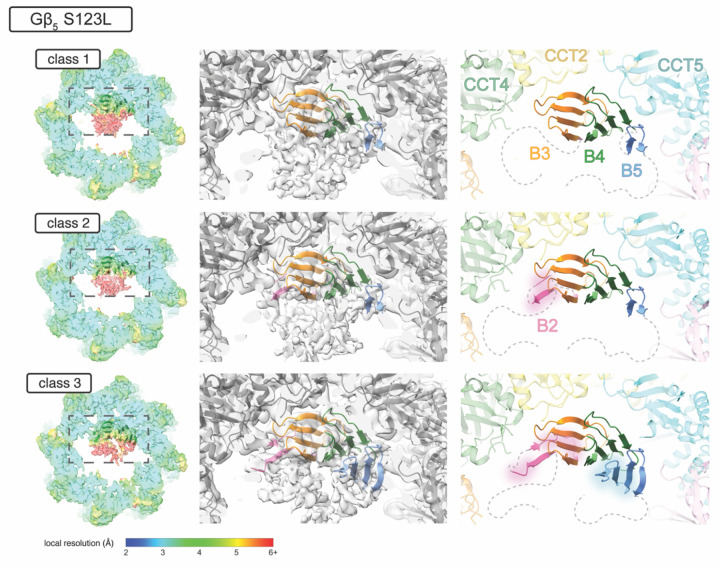
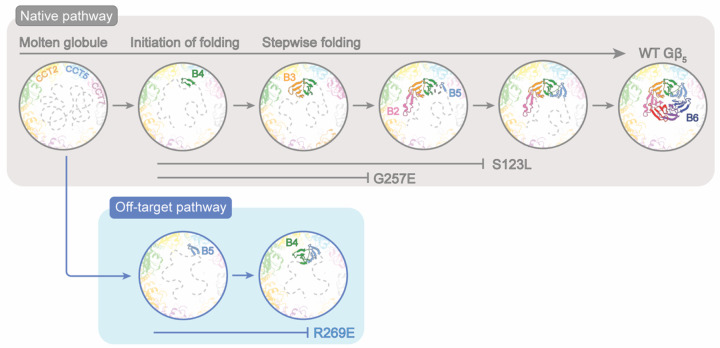
The Chaperonin Containing Tailless polypeptide 1 (CCT or TRiC) is an essential cytosolic chaperone that folds multiple protein substrates, including many with β-propeller folds. One β-propeller substrate is the G protein β5 subunit (Gβ5) of Regulator of G protein Signaling (RGS) complexes that determine the duration of G protein signals in neurons. In recent work, we used cryo-electron microscopy (cryo-EM) to visualize the complete CCT-mediated folding trajectory for Gβ5, from an initiating electrostatic interaction of a single β-strand in Gβ5 with CCT5 to a completely folded β-propeller structure. Here, we used biochemistry and cryo-EM to determine how missense mutations in Gβ5, including those that cause severe neurological diseases, alter the Gβ5 folding trajectory and lead to incompletely folded, trapped intermediates. These findings highlight how defects in chaperonin-mediated folding contribute to disease and suggest potential strategies for stabilizing misfolded proteins to restore function.
Keywords: G protein; chaperonin; cryo-electron microscopy; missense mutations; protein folding.
Conflict of interest statement
Competing Interest Statement: The authors declare no competing interests.
Figures
Similar articles
-
Visualizing the chaperone-mediated folding trajectory of the G protein β5 β-propeller.Mol Cell. 2023 Nov 2;83(21):3852-3868.e6. doi: 10.1016/j.molcel.2023.09.032. Epub 2023 Oct 17. Mol Cell. 2023. PMID: 37852256 Free PMC article.
-
Visualizing the chaperone-mediated folding trajectory of the G protein β5 β-propeller.bioRxiv [Preprint]. 2023 May 4:2023.05.04.539424. doi: 10.1101/2023.05.04.539424. bioRxiv. 2023. Update in: Mol Cell. 2023 Nov 2;83(21):3852-3868.e6. doi: 10.1016/j.molcel.2023.09.032. PMID: 37205387 Free PMC article. Updated. Preprint.
-
Structural visualization of the tubulin folding pathway directed by human chaperonin TRiC/CCT.Cell. 2022 Dec 8;185(25):4770-4787.e20. doi: 10.1016/j.cell.2022.11.014. Cell. 2022. PMID: 36493755 Free PMC article.
-
Immunogenicity and seroefficacy of pneumococcal conjugate vaccines: a systematic review and network meta-analysis.Health Technol Assess. 2024 Jul;28(34):1-109. doi: 10.3310/YWHA3079. Health Technol Assess. 2024. PMID: 39046101 Free PMC article.
-
Signs and symptoms to determine if a patient presenting in primary care or hospital outpatient settings has COVID-19.Cochrane Database Syst Rev. 2022 May 20;5(5):CD013665. doi: 10.1002/14651858.CD013665.pub3. Cochrane Database Syst Rev. 2022. PMID: 35593186 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources