

This is a preprint.
A noncanonical Pol III-dependent, Microprocessor-independent biogenesis pathway generates a germline enriched miRNA family
- PMID: 40501874
- PMCID: PMC12157501
- DOI: 10.1101/2025.04.11.648421
A noncanonical Pol III-dependent, Microprocessor-independent biogenesis pathway generates a germline enriched miRNA family
Update in
-
A noncanonical Pol III-dependent, Microprocessor-independent biogenesis pathway generates a germline-enriched miRNA family.Genes Dev. 2025 Jul 14. doi: 10.1101/gad.352481.124. Online ahead of print. Genes Dev. 2025. PMID: 40659526
Abstract
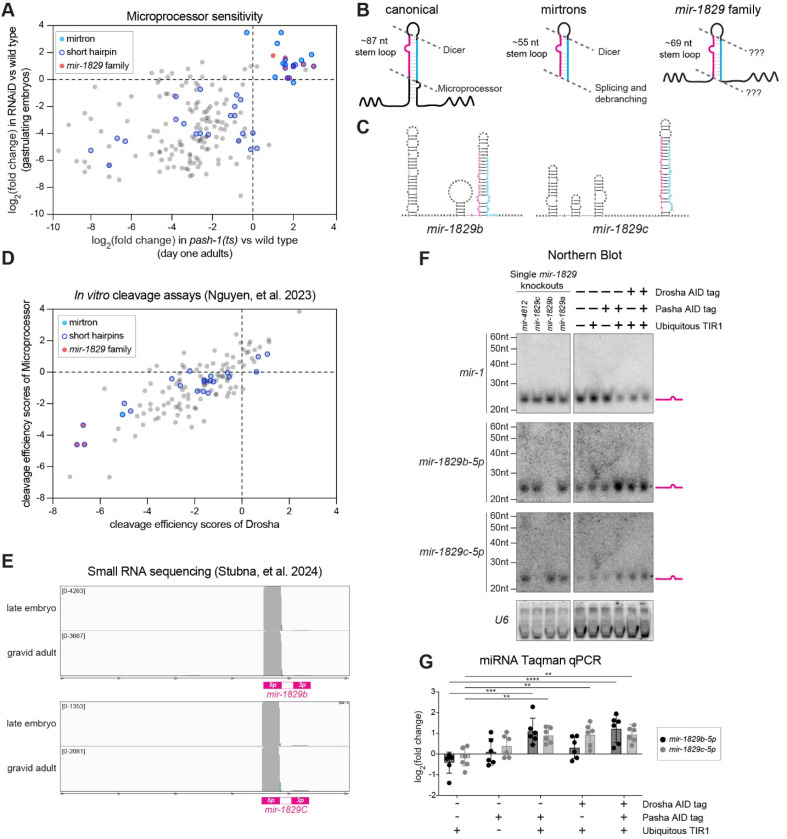
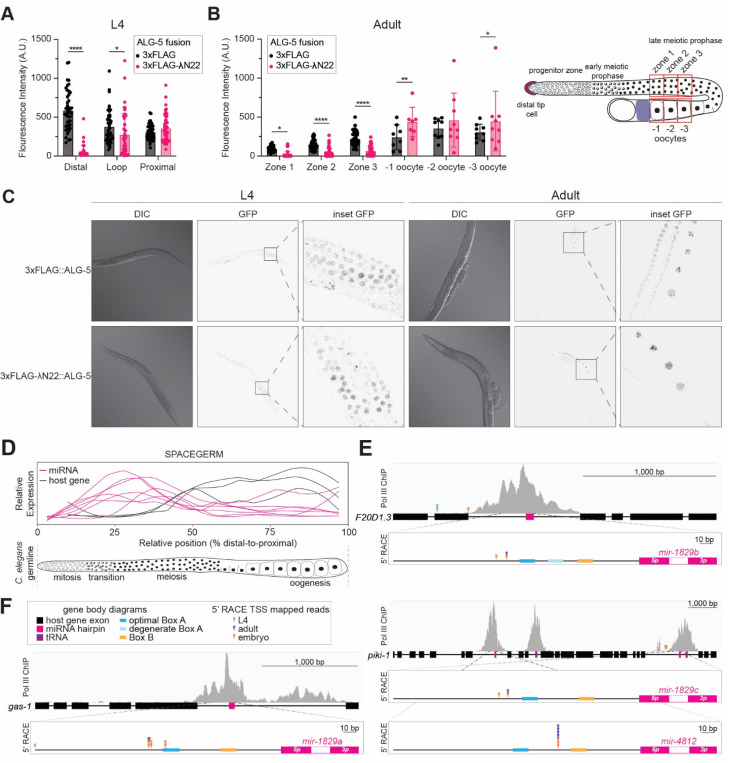
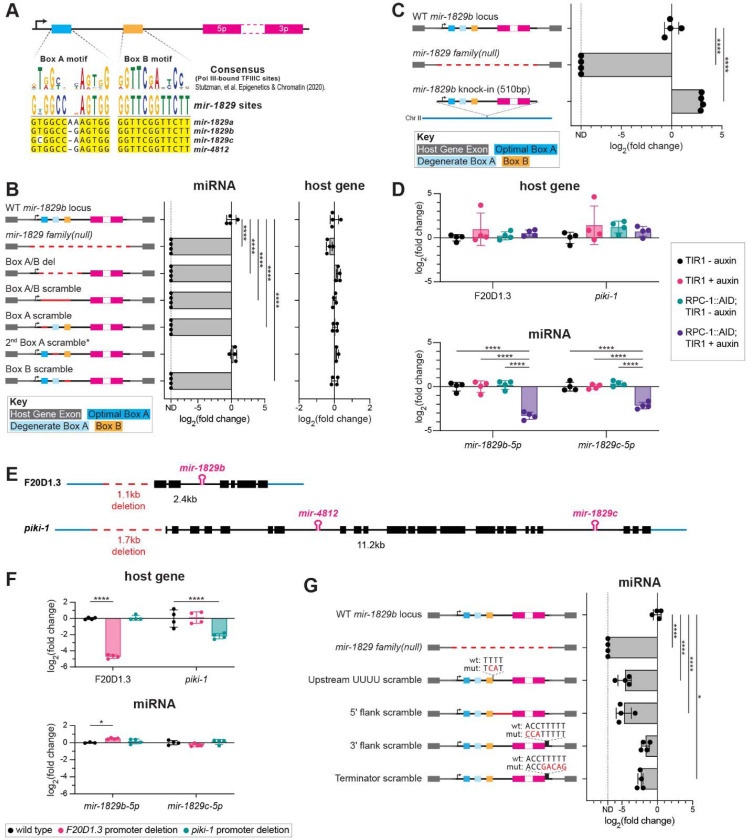
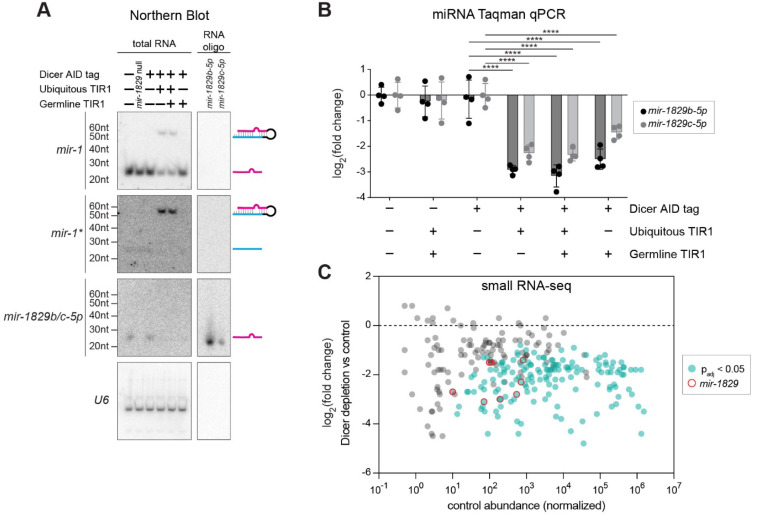
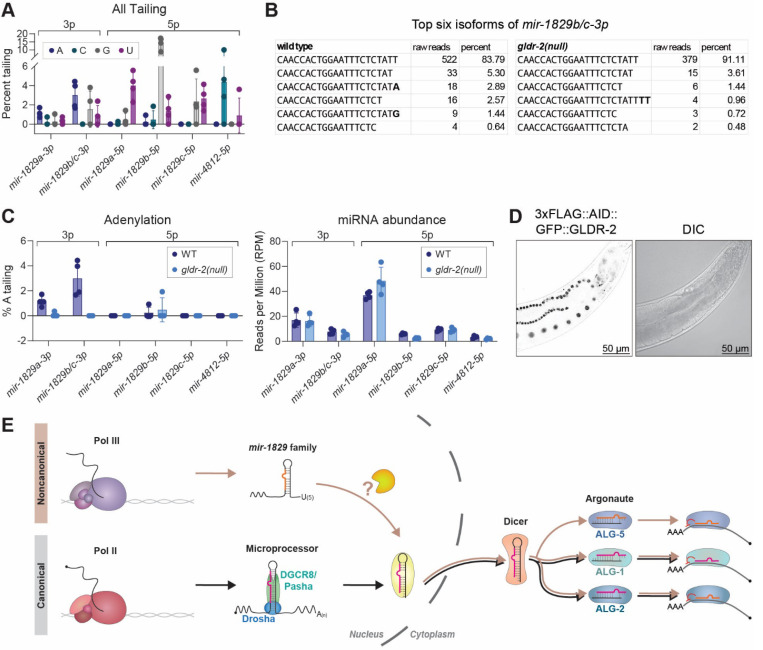
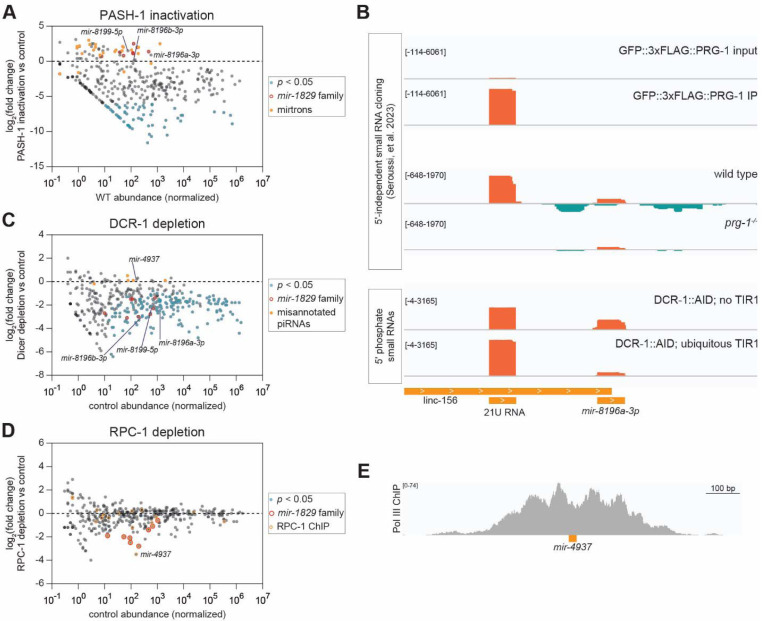
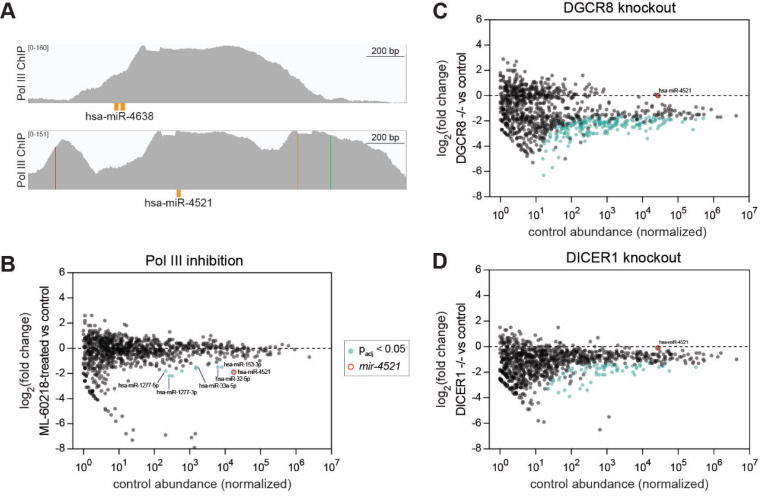
MicroRNAs (miRNAs) are short RNAs that post-transcriptionally regulate gene expression. In canonical miRNA biogenesis, primary miRNAs are transcribed from intergenic loci or intronic regions by RNA polymerase II, sequentially cleaved by the Microprocessor complex and Dicer, and resulting mature miRNAs are loaded into Argonaute to repress target mRNAs. A minority of miRNAs are generated via noncanonical biogenesis pathways that bypass the Microprocessor and/or Dicer. Here, we describe a new Pol III-dependent, Microprocessor-independent, and Dicer-dependent biogenesis pathway exemplified by the mir-1829 family in C. elegans. Although the mir-1829 family loci reside in intronic regions of protein-coding genes, we show that the miRNAs are derived from independent Pol III transcripts. Unlike other Pol III-dependent miRNAs, the mir-1829 family small RNAs are the dominant species derived from their loci, rather than fragments of a larger functional noncoding RNA. These germline-enriched miRNAs are loaded in multiple miRNA Argonautes, including the recently-characterized germline Argonaute ALG-5, which we demonstrate is repressive when tethered to a reporter transcript. We extend these findings, identifying additional Pol III-transcribed and noncanonical small RNAs in C. elegans and human datasets, including human miR-4521. These young, noncanonical miRNAs may represent an early snapshot in the evolution of de novo miRNA genes.
Keywords: DGCR8; Drosha; Pasha; RNA Polymerase III; miR-1277; miR-153; miR-32; miR-33a; mir-8196; pash-1.
Conflict of interest statement
Competing Interest Statement The authors have no competing interests to declare.
Figures
Similar articles
-
A noncanonical Pol III-dependent, Microprocessor-independent biogenesis pathway generates a germline-enriched miRNA family.Genes Dev. 2025 Jul 14. doi: 10.1101/gad.352481.124. Online ahead of print. Genes Dev. 2025. PMID: 40659526
-
Peptidylarginine deiminase 2 regulates expression of DGCR8 affecting miRNA biogenesis in gonadotrope cells.Reproduction. 2023 Jul 3;166(2):125-134. doi: 10.1530/REP-22-0482. Print 2023 Aug 1. Reproduction. 2023. PMID: 37310889 Free PMC article.
-
A MicroRNA Screen Identifies the Wnt Signaling Pathway as a Regulator of the Interferon Response during Flavivirus Infection.J Virol. 2017 Mar 29;91(8):e02388-16. doi: 10.1128/JVI.02388-16. Print 2017 Apr 15. J Virol. 2017. PMID: 28148804 Free PMC article.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
MicroRNAs as biomarkers in spontaneous intracerebral hemorrhage: A systematic review of recent clinical evidence.Clin Neurol Neurosurg. 2022 Feb;213:107130. doi: 10.1016/j.clineuro.2022.107130. Epub 2022 Jan 14. Clin Neurol Neurosurg. 2022. PMID: 35066247
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources