

This is a preprint.
SLX4IP acts in parallel to FANCM to limit BLM-dependent replication stress at ALT telomeres
- PMID: 40501906
- PMCID: PMC12154878
- DOI: 10.1101/2025.05.28.656696
SLX4IP acts in parallel to FANCM to limit BLM-dependent replication stress at ALT telomeres
Abstract
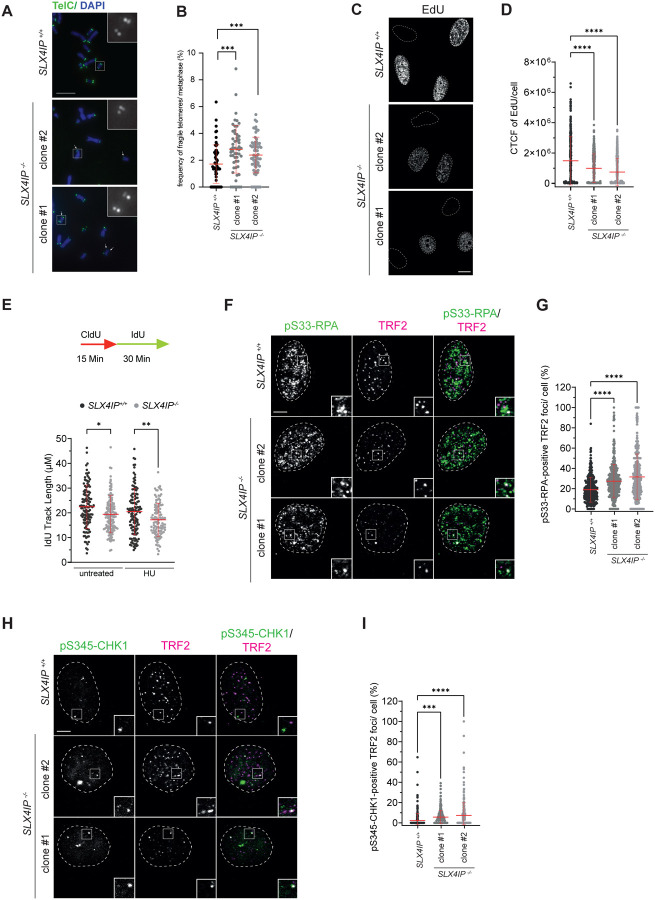
Alternative Lengthening of Telomeres (ALT) is a telomerase-independent telomere maintenance mechanism that enables cancer cells to gain unlimited replicative capacity. ALT relies on recombination-mediated telomere elongation and is promoted by telomeric replication stress. However, ALT requires strict regulation, as excessive replication stress or recombination are cytotoxic. Central to ALT is the RecQ helicase BLM, which regulates telomeric replication stress and promotes telomere recombination and DNA synthesis. Despite its key role in the ALT pathway, BLM must be tightly regulated to prevent deleterious outcomes. Here, we identify SLX4IP as a key suppressor of BLM-driven replication stress at ALT telomeres. Loss of SLX4IP in ALT-positive cells leads to BLM-dependent telomeric replication stress and impaired replication fork progression. Mechanistically, SLX4IP limits the unwinding of unligated Okazaki fragments by BLM on the lagging strand during telomere replication. This reduces the formation of toxic 5' DNA flaps and prevents hyperactivation of ATR signalling and deleterious recombination levels. We also uncover a synthetic lethal interaction between SLX4IP and FANCM, an ATPase/translocase that is a known regulator of BLM at telomeric replication forks in ALT cells. We demonstrate that SLX4IP and FANCM act in parallel to restrain BLM activity, thereby maintaining the balance of replication stress and recombination that is necessary for productive ALT. These findings reveal a vulnerability in ALT-positive cancers lacking SLX4IP and establish SLX4IP as a potential biomarker for therapeutic strategies targeting FANCM.
Keywords: ALT; BLM; FANCM; SLX4IP; cancer; genome stability; lagging-strand replication; replication stress; telomere.
Conflict of interest statement
DECLARATION OF INTERESTS The authors declare no completing interests.
Figures



References
-
- Bryan T. M., Englezou A., Dalla-Pozza L., Dunham M. A. & Reddel R. R. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat Med 3, 1271–1274 (1997). - PubMed
-
- Henson J. D. & Reddel R. R. Assaying and investigating Alternative Lengthening of Telomeres activity in human cells and cancers. FEBS Letters 584, 3800–3811 (2010). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous