

This is a preprint.
Structural basis of liver de-targeting and neuronal tropism of CNS-targeted AAV capsids
- PMID: 40501988
- PMCID: PMC12157541
- DOI: 10.1101/2025.06.02.655683
Structural basis of liver de-targeting and neuronal tropism of CNS-targeted AAV capsids
Abstract
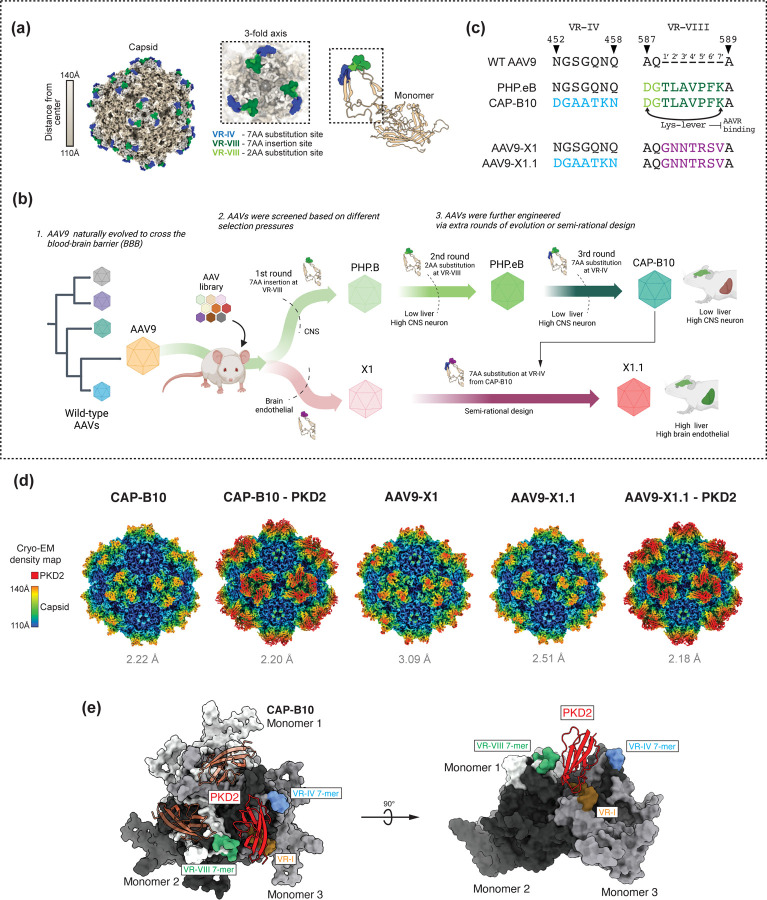
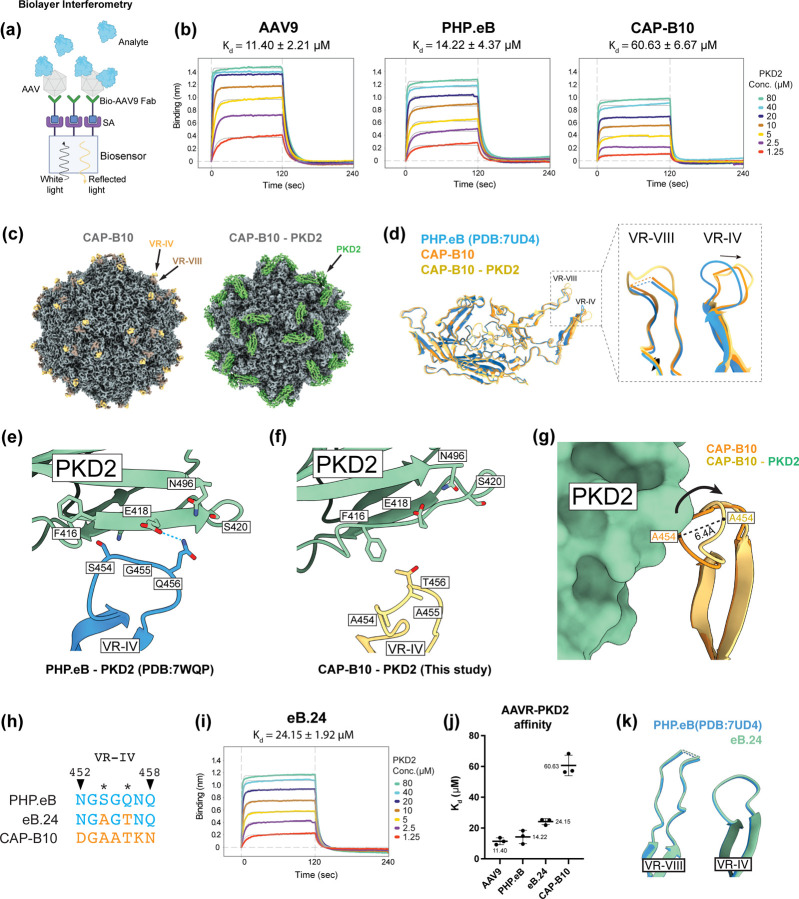
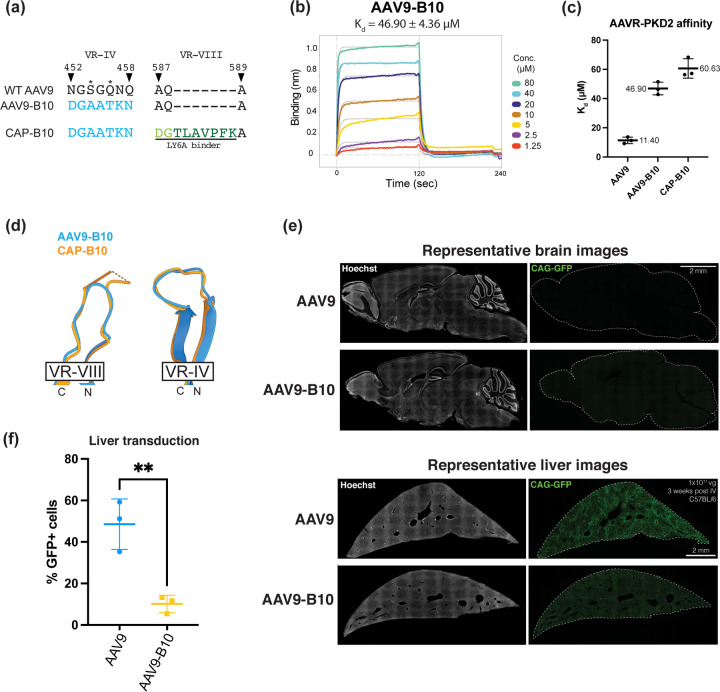
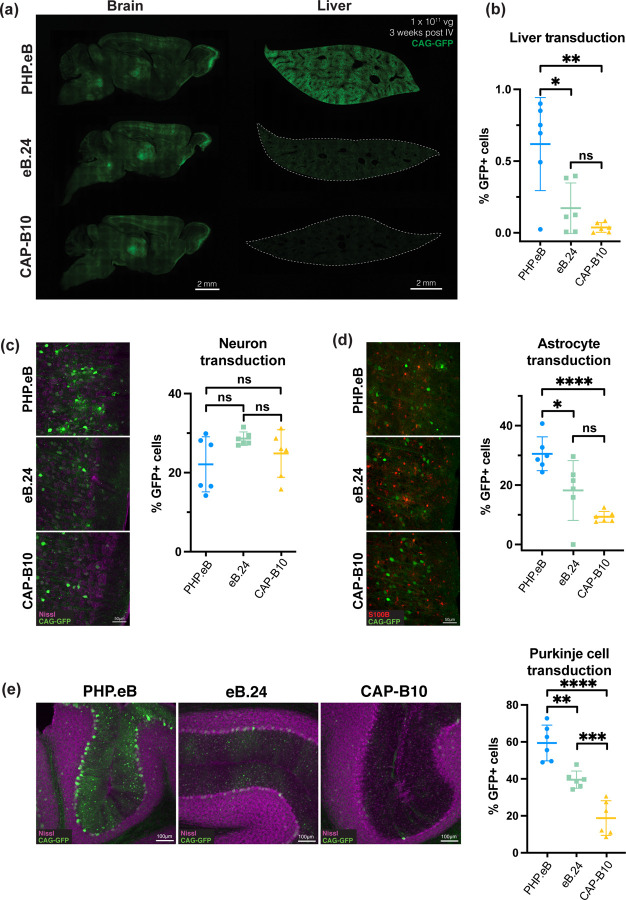
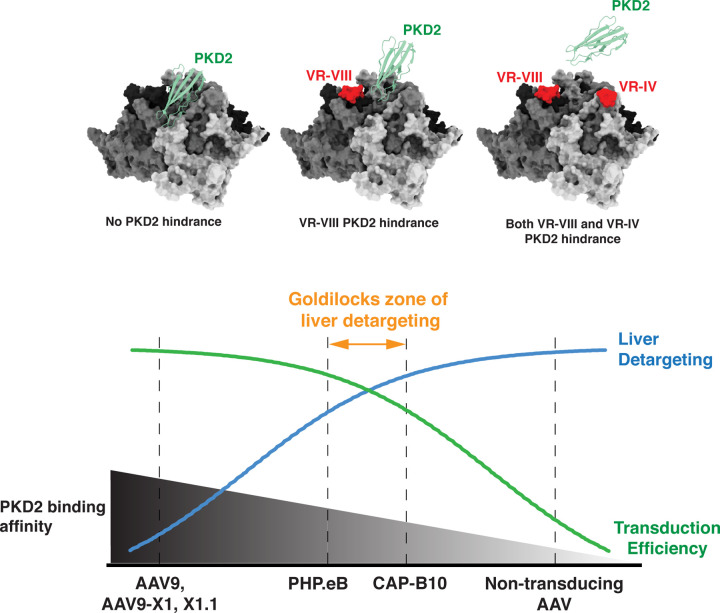
Crossing the blood-brain barrier while minimizing liver transduction is a key challenge in developing safe adeno-associated virus (AAV) vectors for treating brain disorders. In mice, the engineered capsid PHP.eB shows enhanced brain transduction, while the further engineered CAP-B10 is also de-targeted from astrocytes and liver. Here, we solve cryo-EM structures of CAP-B10 and its complex with AAV receptor (AAVR) domain PKD2, at 2.22 and 2.20 Å resolutions, respectively. These structures reveal a structural motif that hinders AAVR binding, which we confirm by measuring affinities. We show that this motif is transferable to other capsids by solving cryo-EM structures of AAV9-X1 and AAV9-X1.1, without and with PKD2, at 3.09, 2.51, and 2.18 Å, respectively. Using this structural information, we designed and validated novel AAV variants with reduced liver and altered brain cell tropism in vivo. Overall, our findings demonstrate that rationally modulating AAVR affinity can alter liver targeting and cellular tropism.
Conflict of interest statement
Competing interests V.G. is a cofounder and board member of Capsida Biotherapeutics, a fully integrated AAV engineering and gene therapy company. The remaining authors declare no competing interests.
Figures

References
-
- Ling Q., Herstine J. A., Bradbury A. & Gray S. J. AAV-based in vivo gene therapy for neurological disorders. Nat. Rev. Drug Discov. 22, 789–806 (2023). - PubMed
-
- Al-Zaidy S. A. & Mendell J. R. From Clinical Trials to Clinical Practice: Practical Considerations for Gene Replacement Therapy in SMA Type 1. Pediatr. Neurol. 100, 3–11 (2019). - PubMed
-
- Kang L. et al. AAV vectors applied to the treatment of CNS disorders: Clinical status and challenges. J. Controlled Release 355, 458–473 (2023). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous