

This is a preprint.
Transposon-plasmid nesting enables fast response to fluctuating environments
- PMID: 40502034
- PMCID: PMC12157364
- DOI: 10.1101/2025.06.04.657954
Transposon-plasmid nesting enables fast response to fluctuating environments
Abstract
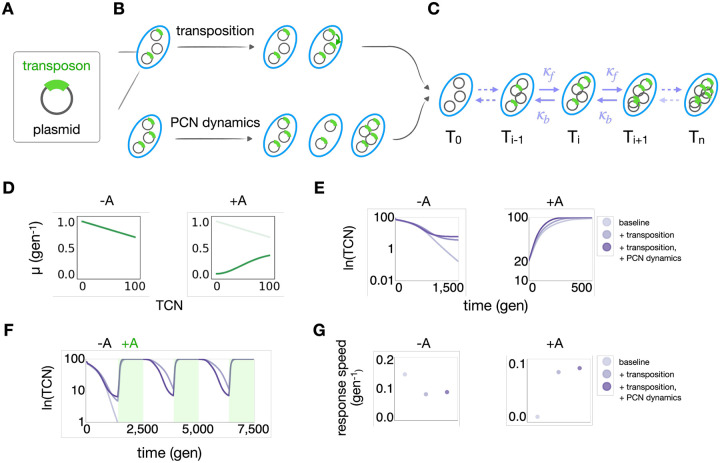
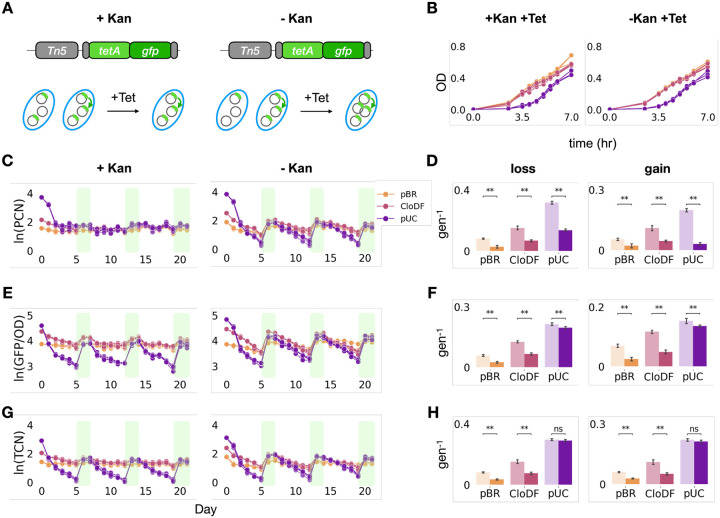
Mobile genetic elements (MGEs) play a critical role in shaping the response and evolution of microbial populations and communities. Despite distinct maintenance mechanisms, different types of MGEs can form nested structures. Using bioinformatics analysis of 14,338 plasmids in the NCBI RefSeq database, we found transposons to be widespread and significantly enriched on plasmids relative to chromosomes, highlighting the prevalence of transposon-plasmid nesting. We hypothesized that this nested structure provides unique adaptive advantages by combining transposition-driven genetic mobility with plasmid-mediated copy number amplification. Using engineered transposon systems, we demonstrated that nesting enables rapid and tunable responses of transposon-encoded genes in fluctuating environments. Specifically, transposition maintains a reservoir of the encoded genes, while plasmid copy number fluctuations further amplify the dynamic range of gene dosage, thus enhancing the response speed and stability of transposon-encoded traits. Our findings demonstrate an adaptive benefit of transposon-plasmid nesting and provide insights into their ecological persistence and evolutionary success.
Conflict of interest statement
Disclosure and competing interests statement The authors declare that they have no conflict of interest.
Figures



Similar articles
-
Transposon-directed insertion-site sequencing (TraDIS) analysis of Enterococcus faecium using nanopore sequencing and a WebAssembly analysis platform.Microbiol Spectr. 2025 Jul;13(7):e0062825. doi: 10.1128/spectrum.00628-25. Epub 2025 Jun 10. Microbiol Spectr. 2025. PMID: 40494643 Free PMC article.
-
Mechanistic divergence between SOS response activation and antibiotic-induced plasmid conjugation in Escherichia coli.Microbiol Spectr. 2025 Jul;13(7):e0009025. doi: 10.1128/spectrum.00090-25. Epub 2025 May 28. Microbiol Spectr. 2025. PMID: 40434128 Free PMC article.
-
Effectiveness and cost-effectiveness of computer and other electronic aids for smoking cessation: a systematic review and network meta-analysis.Health Technol Assess. 2012;16(38):1-205, iii-v. doi: 10.3310/hta16380. Health Technol Assess. 2012. PMID: 23046909
-
Accreditation through the eyes of nurse managers: an infinite staircase or a phenomenon that evaporates like water.J Health Organ Manag. 2025 Jun 30. doi: 10.1108/JHOM-01-2025-0029. Online ahead of print. J Health Organ Manag. 2025. PMID: 40574247
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
Cited by
-
Protease activated receptor inhibitors in rheumatoid arthritis: a new frontier in treatment.Inflammopharmacology. 2025 Jul;33(7):3583-3595. doi: 10.1007/s10787-025-01822-x. Epub 2025 Jun 28. Inflammopharmacology. 2025. PMID: 40580298 Review.
References
-
- Roy T., Bandopadhyay A., Paul C., Majumdar S., & Das N. (2022). Role of plasmid in pesticide degradation and metal tolerance in two plant growth-promoting Rhizobacteria bacillus cereus (NCIM 5557) and Bacillus safensis (NCIM 5558). Current Microbiology, 79(4). 10.1007/s00284-022-02793-w - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources