

This is a preprint.
Identification of cross-stage, cross-species malaria CD8+ T cell antigens
- PMID: 40502747
- PMCID: PMC12154130
- DOI: 10.21203/rs.3.rs-6682089/v1
Identification of cross-stage, cross-species malaria CD8+ T cell antigens
Abstract
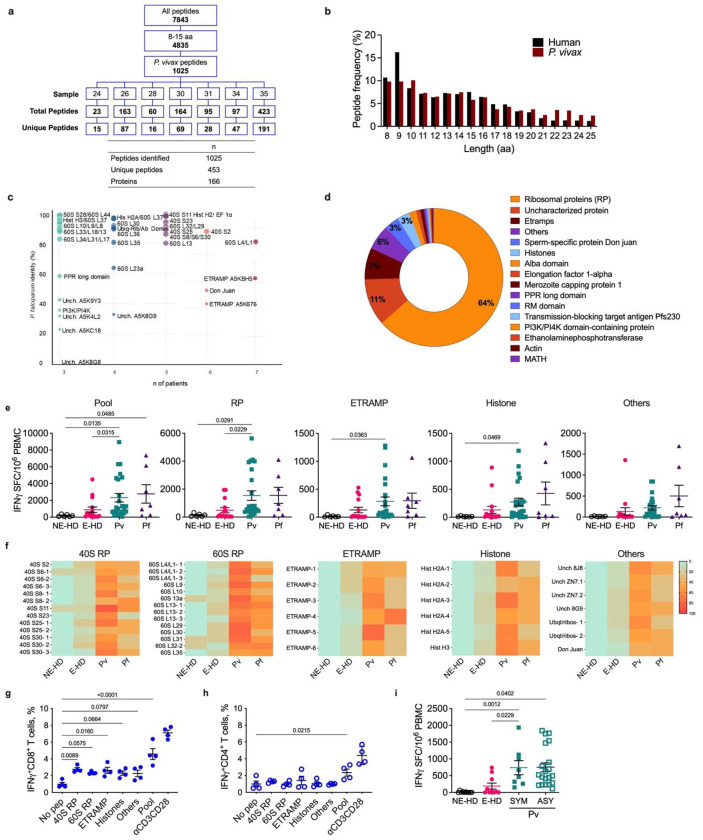
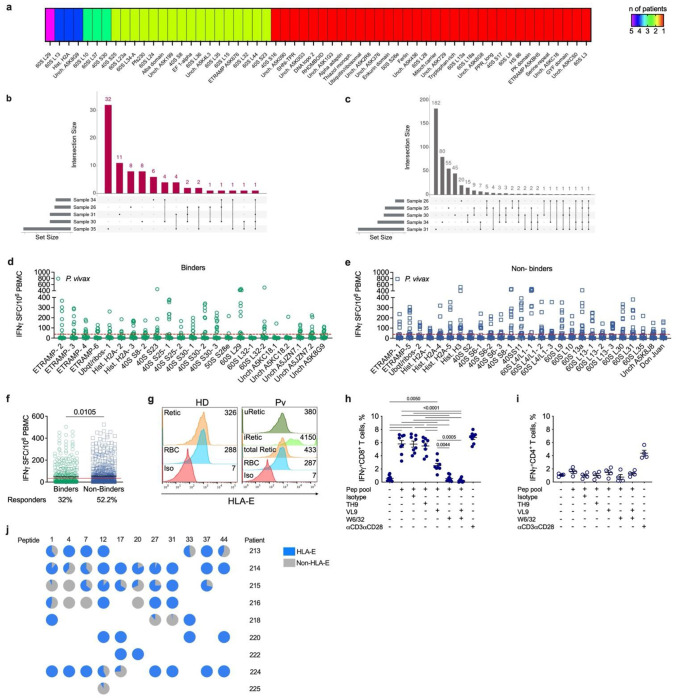
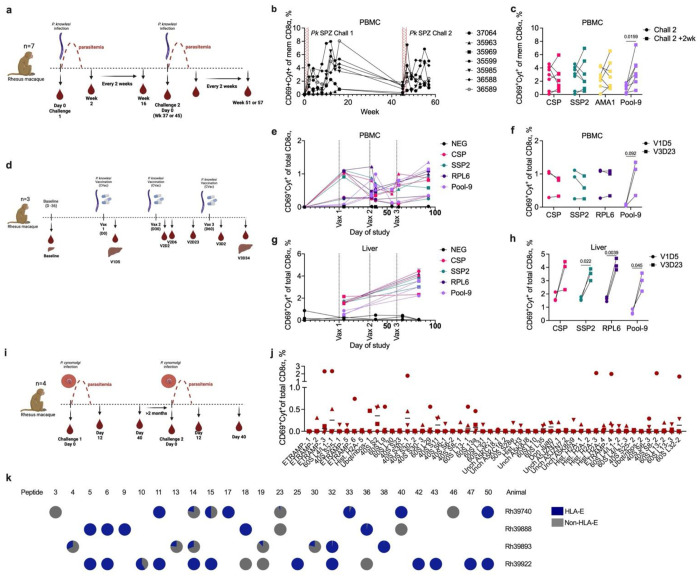
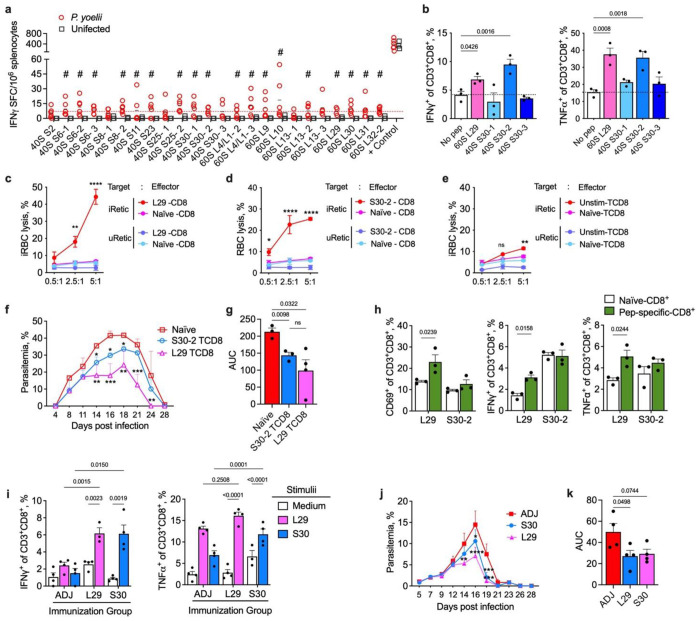
Malaria is one of the most prevalent parasitic diseases in the world. In 2023, 263 million malaria cases were estimated worldwide1. Two species of Plasmodium, P. falciparum and P. vivax, cause most human malaria. Despite the licensing of two partially protective vaccines for P. falciparum, there is no vaccine capable of providing long-term control or elimination2,3. A major limitation for vaccine development is the lack of validated T cell epitopes for either species that could be targeted by vaccines. P. vivax is the most widespread human malaria parasite and is the major species causing malaria in the Americas and Asia while P. falciparum is more prevalent in Africa1. P. vivax exclusively infects reticulocytes in peripheral blood, which, unlike the mature erythrocytes infected by P. falciparum, still retain RNA and therefore retain host protein translation capabilities4. We previously reported that P. vivax-infected reticulocytes express the major human leukocyte antigen class I (HLA-I), which allows parasite sensing by CD8+ T cells and consequent killing of parasite-infected host cells and intracellular parasites5. Here we report by immunopeptidomic analysis the first unbiased identification of Plasmodium spp. antigens presented via HLA-I on infected reticulocytes. We identified 453 unique peptides that mapped to 166 different proteins. Most of these antigens were housekeeping proteins that are constitutively expressed at multiple stages of the parasite life cycle. Common peptides were presented in different individuals by the same or distinct HLA-ABC alleles as well as by non-classical HLA-E. Many peptide sequences were highly conserved in P. falciparum and P. vivax. The immunogenicity of the newly identified epitopes was validated in both P. vivax- and P. falciparum-infected patient samples. Furthermore, several of these antigens were immunogenic in the blood and liver of non-human primates following Plasmodium infection and attenuated parasite immunization. Two antigens were also the target of protective CD8+ T cell-mediated immunity in rodents. Thus, these antigens have potential for use in a cross-stage and cross-species malaria vaccine.
Conflict of interest statement
Additional Declarations: There is NO Competing Interest. Competing interest declaration The authors declare no competing financial interests.
Figures
References
-
- WHO. World malaria report 2024. (World Health Organization, Geneva, 2024).
METHODS ONLY REFERENCES
-
- Boyum A. Isolation of mononuclear cells and granulocytes from human blood. Isolation of monuclear cells by one centrifugation, and of granulocytes by combining centrifugation and sedimentation at 1 g. Scand J Clin Lab Invest Suppl 97, 77–89 (1968). - PubMed
-
- The Plasmodium Genome Database, C. PlasmoDB: An integrative database of the Plasmodium falciparum genome. Tools for accessing and analyzing finished and unfinished sequence data. The Plasmodium Genome Database Collaborative. Nucleic Acids Res 29, 66–69 (2001). 10.1093/nar/29.1.66 - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
