

Global siRNA screen identifies human host factors critical for SARS-CoV-2 replication and late stages of infection
- PMID: 40504864
- PMCID: PMC12204624
- DOI: 10.1371/journal.pbio.3002738
Global siRNA screen identifies human host factors critical for SARS-CoV-2 replication and late stages of infection
Abstract
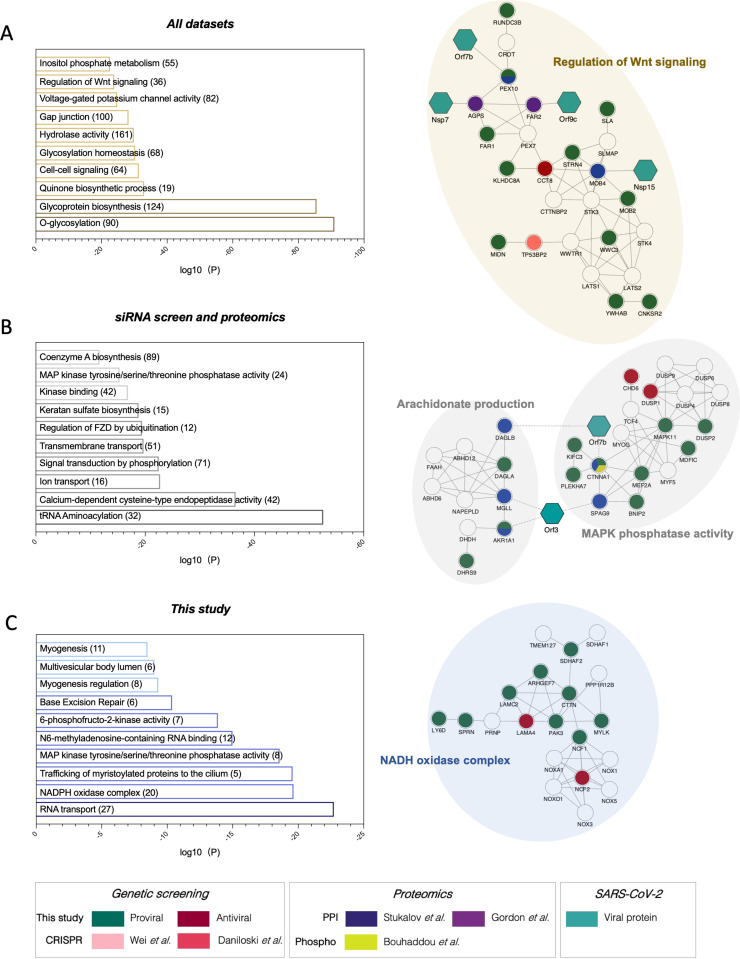
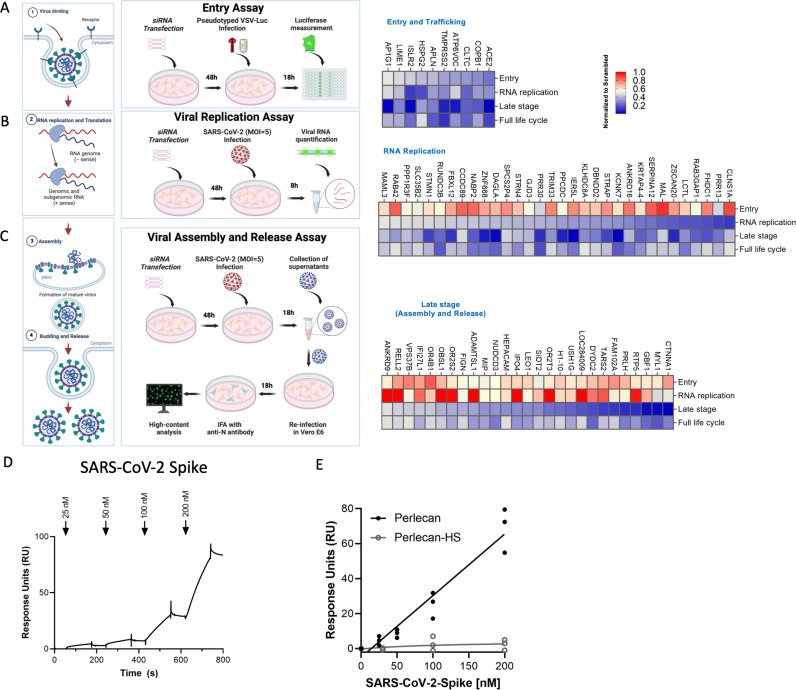
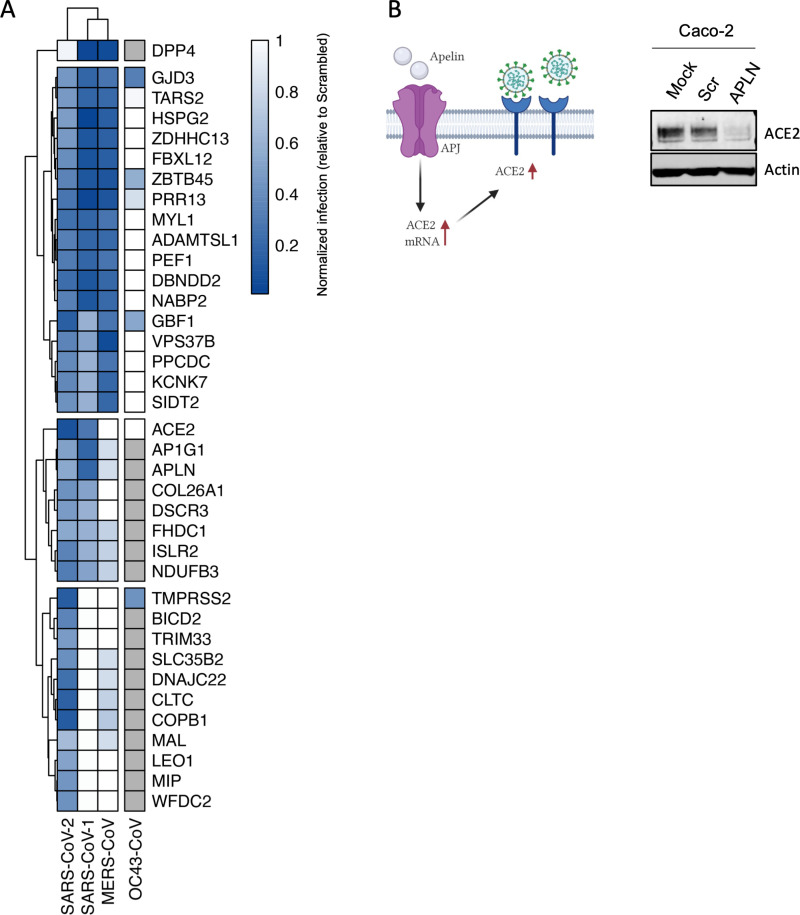
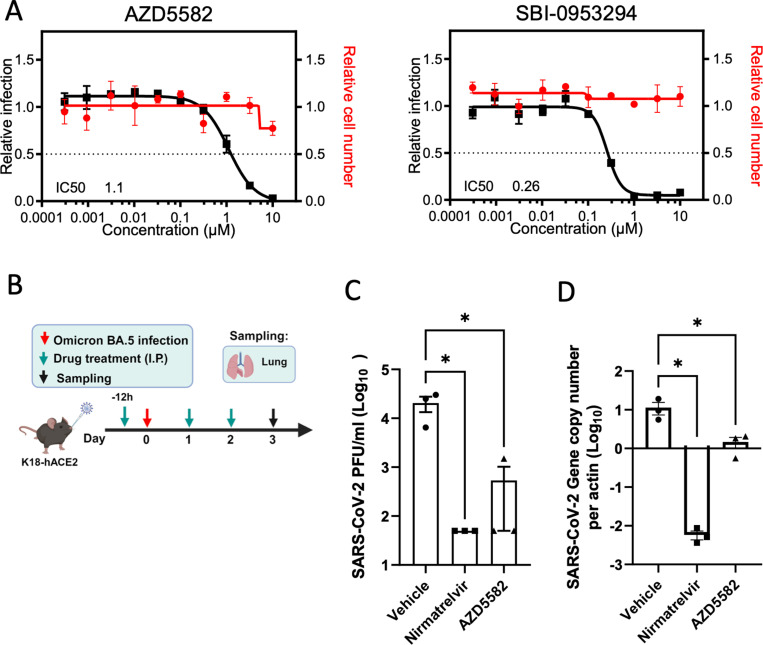
Defining the subset of cellular factors governing SARS-CoV-2 replication can provide critical insights into viral pathogenesis and identify targets for host-directed antiviral therapies. While a number of genetic screens have previously reported SARS-CoV-2 host dependency factors, most of these approaches relied on utilizing pooled genome-scale CRISPR libraries, which are biased toward the discovery of host proteins impacting early stages of viral replication. To identify host factors involved throughout the SARS-CoV-2 infectious cycle, we conducted an arrayed genome-scale siRNA screen. Resulting data were integrated with published functional screens and proteomics data to reveal (i) common pathways that were identified in all OMICs datasets-including regulation of Wnt signaling and gap junctions, (ii) pathways uniquely identified in this screen-including NADH oxidation, or (iii) pathways supported by this screen and proteomics data but not published functional screens-including arachionate production and MAPK signaling. The identified proviral host factors were mapped into the SARS-CoV-2 infectious cycle, including 32 proteins that were determined to impact viral replication and 27 impacting late stages of infection, respectively. Additionally, a subset of proteins was tested across other coronaviruses revealing a subset of proviral factors that were conserved across pandemic SARS-CoV-2, epidemic SARS-CoV-1 and MERS-CoV, and the seasonal coronavirus OC43-CoV. Further studies illuminated a role for the heparan sulfate proteoglycan perlecan in SARS-CoV-2 viral entry and found that inhibition of the non-canonical NF-kB pathway through targeting of BIRC2 restricts SARS-CoV-2 replication both in vitro and in vivo. These studies provide critical insight into the landscape of virus-host interactions driving SARS-CoV-2 replication as well as valuable targets for host-directed antivirals.
Copyright: © 2025 Yin et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: J.F.H. has received research support, paid to Northwestern University, from Gilead Sciences, and is a paid consultant for Merck. The A.G.-S. laboratory has received research support from GSK, Pfizer, Senhwa Biosciences, Kenall Manufacturing, Blade Therapeutics, Avimex, Johnson & Johnson, Dynavax, 7Hills Pharma, Pharmamar, ImmunityBio, Accurius, Nanocomposix, Hexamer, N-fold LLC, Model Medicines, Atea Pharma, Applied Biological Laboratories and Merck, outside of the reported work. A.G.-S. has consulting agreements for the following companies involving cash and/or stock: Castlevax, Amovir, Vivaldi Biosciences, Contrafect, 7Hills Pharma, Avimex, Pagoda, Accurius, Esperovax, Applied Biological Laboratories, Pharmamar, CureLab Oncology, CureLab Veterinary, Synairgen, Paratus, Pfizer and Prosetta, outside of the reported work. A.G.-S. has been an invited speaker in meeting events organized by Seqirus, Janssen, Abbott, Astrazeneca, and Novavax. A.G.-S. is inventor on patents and patent applications on the use of antivirals and vaccines for the treatment and prevention of virus infections and cancer, owned by the Icahn School of Medicine at Mount Sinai, New York, outside of the reported work. T.I. is cofounder of Data4Cure, is on the Scientific Advisory Board, and has an equity interest. T.I. is on the Scientific Advisory Board of Ideaya BioSciences and has an equity interest. The terms of these arrangements have been reviewed and approved by the University of California San Diego, in accordance with its conflict of interest policies. All other authors declare no competing interests.
Figures

Update of
-
Global siRNA Screen Reveals Critical Human Host Factors of SARS-CoV-2 Multicycle Replication.bioRxiv [Preprint]. 2024 Jul 10:2024.07.10.602835. doi: 10.1101/2024.07.10.602835. bioRxiv. 2024. Update in: PLoS Biol. 2025 Jun 12;23(6):e3002738. doi: 10.1371/journal.pbio.3002738. PMID: 39026801 Free PMC article. Updated. Preprint.
References
-
- The evolution of SARS-CoV-2 | Nature Reviews Microbiology https://www.nature.com/articles/s41579-023-00878-2. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
