

Abscisic acid enhances DNA damage response through the nuclear shuttling of clathrin light chain 2 in plant cells
- PMID: 40512838
- PMCID: PMC12164954
- DOI: 10.1126/sciadv.adt2842
Abscisic acid enhances DNA damage response through the nuclear shuttling of clathrin light chain 2 in plant cells
Abstract
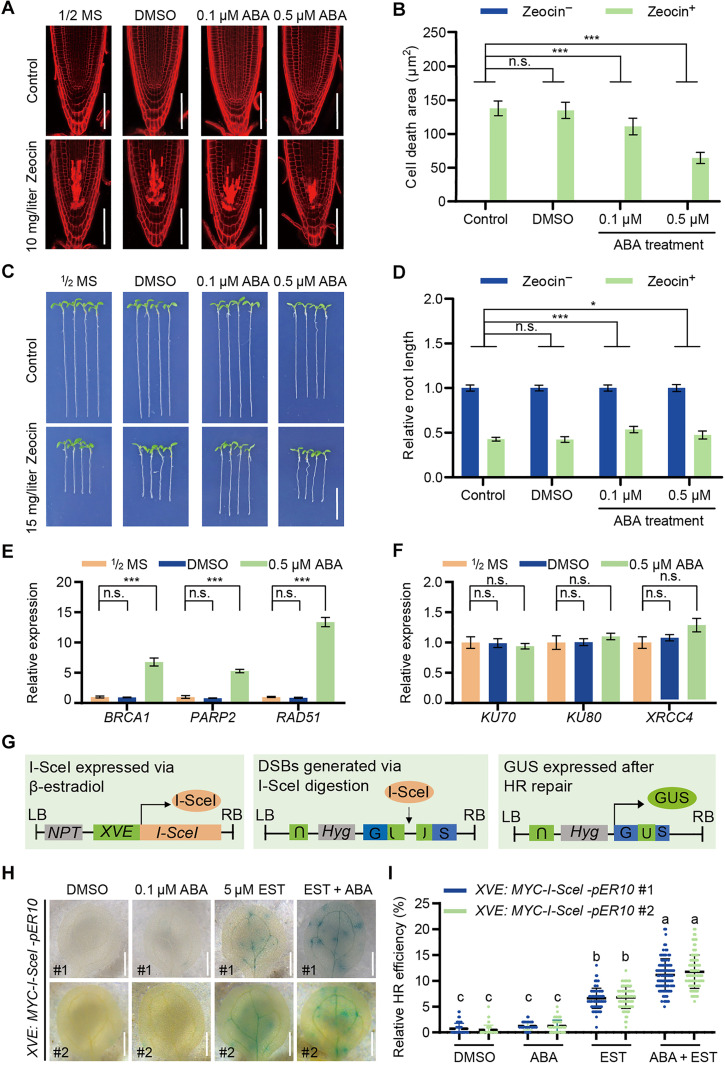
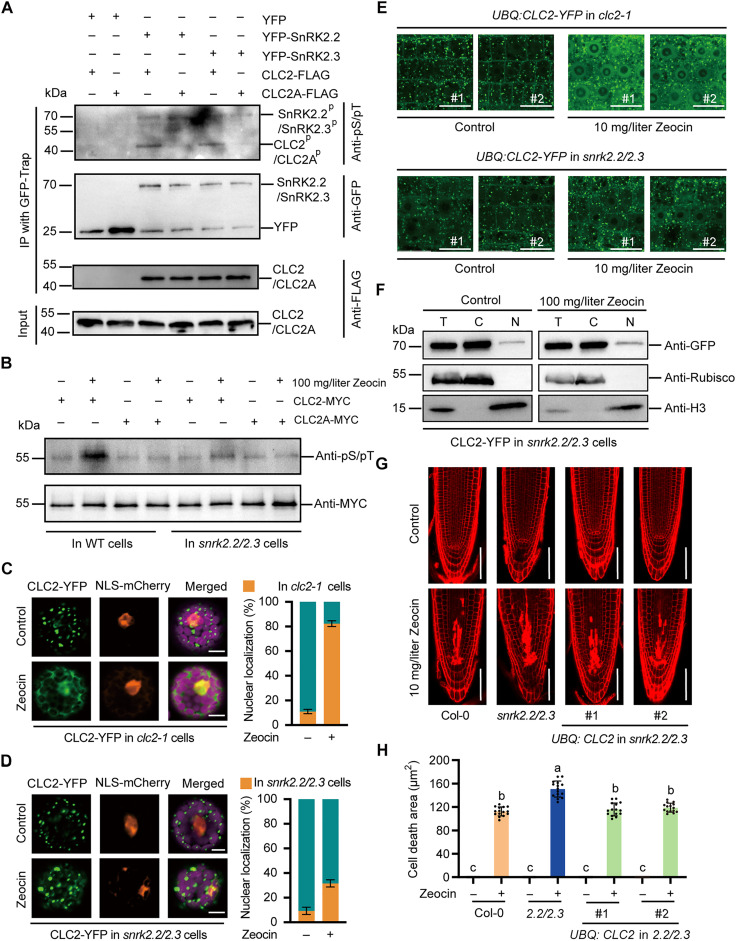
DNA damage arises from various environmental stresses, and ABA is well known for its roles in plant stress resistance. However, its function in plant DNA damage tolerance remains unclear. In this study, we showed that ABA supplementation significantly enhances plant tolerance to DNA-damaging treatments. SnRK2.2 and SnRK2.3 kinases in the ABA signaling pathway are pivotal in this process. These kinases interact with clathrin light chain 2 (CLC2), facilitating its phosphorylation and nuclear translocation in response to Zeocin and ABA treatment. In the nucleus, CLC2 interacts with ADA2b, an adaptor protein crucial for recruiting SMC5/6 complex to the double-strand break (DSB) sites. The enhanced nuclear localization of CLC2 is essential for the accurate localization of ADA2b at DSB sites. Collectively, our study uncovers that ABA enhances plant DNA damage tolerance with a distinct function of CLC2 in genomic stability maintaining, thereby improving our understanding of DNA damage tolerance mechanisms in plants.
Figures




Similar articles
-
CLC2 (Clathrin Light Chain 2)-ATG8h/ATG8i interactions connect clathrin-mediated endocytosis (CME) and the autophagy pathway.Autophagy. 2025 Jan;21(1):246-248. doi: 10.1080/15548627.2024.2414451. Epub 2024 Oct 16. Autophagy. 2025. PMID: 39396122
-
Overexpression of MEKK18 from Arabidopsis pumila in rice significantly enhances stress resistance at the early stage.PLoS One. 2025 Jun 25;20(6):e0325550. doi: 10.1371/journal.pone.0325550. eCollection 2025. PLoS One. 2025. PMID: 40561074 Free PMC article.
-
Abscisic acid enhances non-photochemical quenching through SnRK2 and ABI3 in Physcomitrium patens.J Plant Res. 2025 Jul;138(4):625-636. doi: 10.1007/s10265-025-01627-7. Epub 2025 Apr 7. J Plant Res. 2025. PMID: 40192897
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2021 Apr 19;4(4):CD011535. doi: 10.1002/14651858.CD011535.pub4. Cochrane Database Syst Rev. 2021. Update in: Cochrane Database Syst Rev. 2022 May 23;5:CD011535. doi: 10.1002/14651858.CD011535.pub5. PMID: 33871055 Free PMC article. Updated.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
References
-
- Todorova T., Miteva D., Chankova S., DNA damaging effect of zeocin and methyl methanesulfonate in saccharomyces cerevisiae measured by CFGE. C. R. Acad. Bulg. Sci. 68, 71–78 (2015).
-
- Pedroza-Garcia J. A., Xiang Y., De Veylder L., Cell cycle checkpoint control in response to DNA damage by environmental stresses. Plant J. 109, 490–507 (2022). - PubMed
-
- Moorhead G. B. G., Trinkle-Mulcahy L., Ulke-Lemée A., Emerging roles of nuclear protein phosphatases. Nat. Rev. Mol. Cell Biol. 8, 234–244 (2007). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
