

Switchable client specificity in a dual functional chaperone coordinates light-harvesting complex biogenesis
- PMID: 40512866
- PMCID: PMC12164976
- DOI: 10.1126/sciadv.adu5791
Switchable client specificity in a dual functional chaperone coordinates light-harvesting complex biogenesis
Abstract
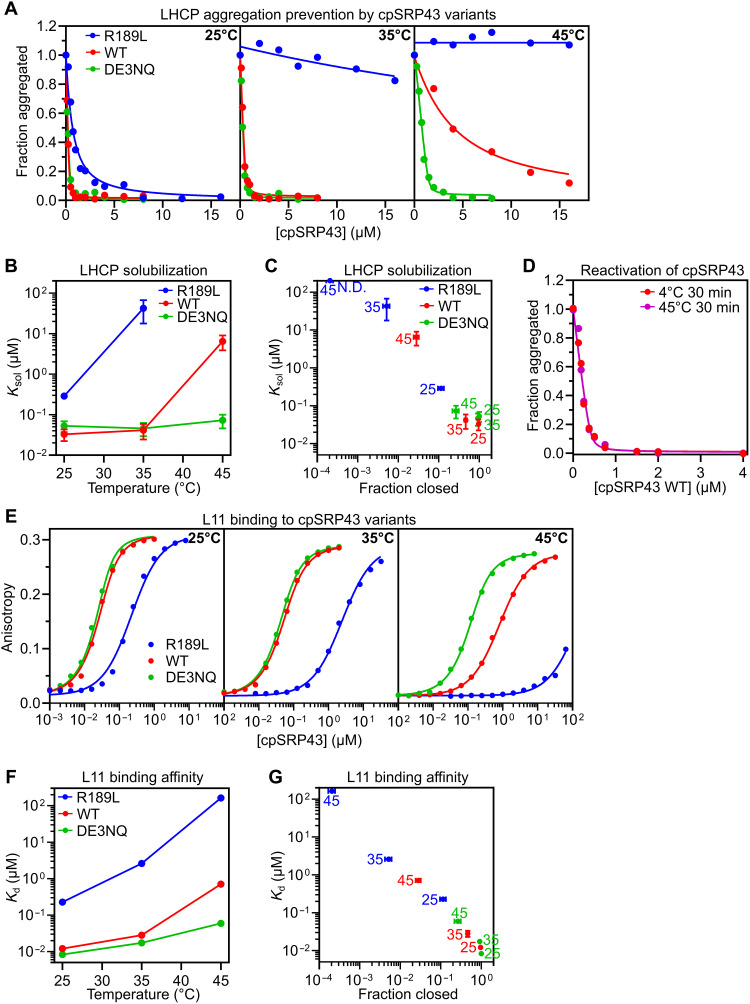
The proper assembly of light-harvesting complexes (LHCs) is critical for photosynthesis and requires the biogenesis of light-harvesting chlorophyll a,b-binding proteins (LHCPs) to be coordinated with chlorophyll (Chl) biosynthesis. The mechanism underlying this coordination is not well understood. Here, we show that a conserved molecular chaperone, chloroplast signal recognition particle 43-kDa protein (cpSRP43), provides a molecular thermostat that helps maintain this coordination. cpSRP43 undergoes a conformational rearrangement between a well-folded closed state and a partially disordered open state. Closed cpSRP43 is dedicated to the biogenesis of LHCPs, whereas open cpSRP43 protects multiple Chl biosynthesis enzymes from heat-induced destabilization. Rising temperature shifts cpSRP43 to the open state, enabling it to protect heat-destabilized Chl biosynthesis enzymes. Our results reveal the molecular basis of a posttranslational mechanism for the thermoadaptation of LHC biogenesis. They also demonstrate how an adenosine triphosphate-independent chaperone uses conformational dynamics to switch its activity and client selectivity, thereby adapting to different proteostatic demands under shifting environmental conditions.
Figures






References
-
- Jarvis P., López-Juez E., Biogenesis and homeostasis of chloroplasts and other plastids. Nat. Rev. Mol. Cell Biol. 14, 787–802 (2013). - PubMed
-
- Dall’Osto L., Bressan M., Bassi R., Biogenesis of light harvesting proteins. Biochim. Biophys. Acta Bioenerg. 1847, 861–871 (2015). - PubMed
-
- Paulsen H., Finkenzeller B., Kühlein N., Pigments induce folding of light-harvesting chlorophyll a/b-binding protein. Eur. J. Biochem. 215, 809–816 (1993). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
