

Disruption of the biorhythm in gastric epithelial cell triggers inflammation in Helicobacter pylori-associated gastritis by aberrantly regulating NFIL3 via CagA activated ERK-SP1 pathway
- PMID: 40518541
- PMCID: PMC12168321
- DOI: 10.1186/s12964-025-02302-z
Disruption of the biorhythm in gastric epithelial cell triggers inflammation in Helicobacter pylori-associated gastritis by aberrantly regulating NFIL3 via CagA activated ERK-SP1 pathway
Abstract
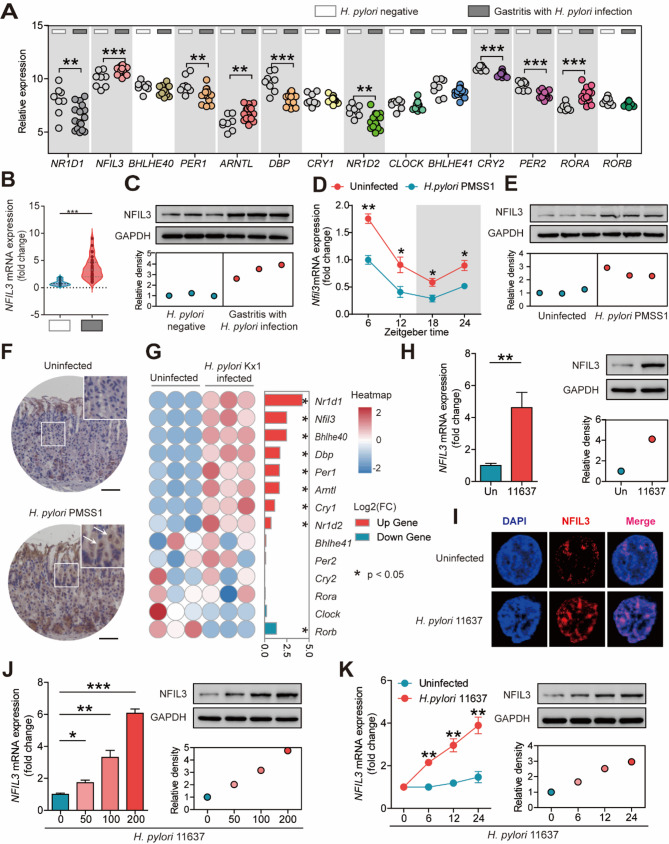
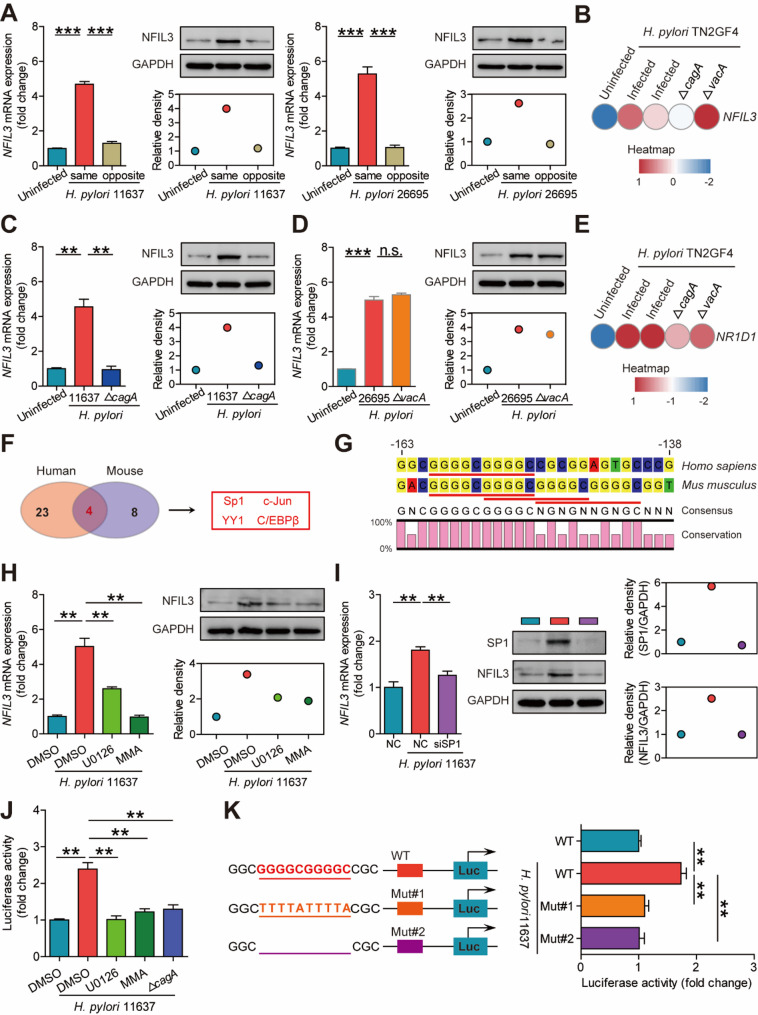
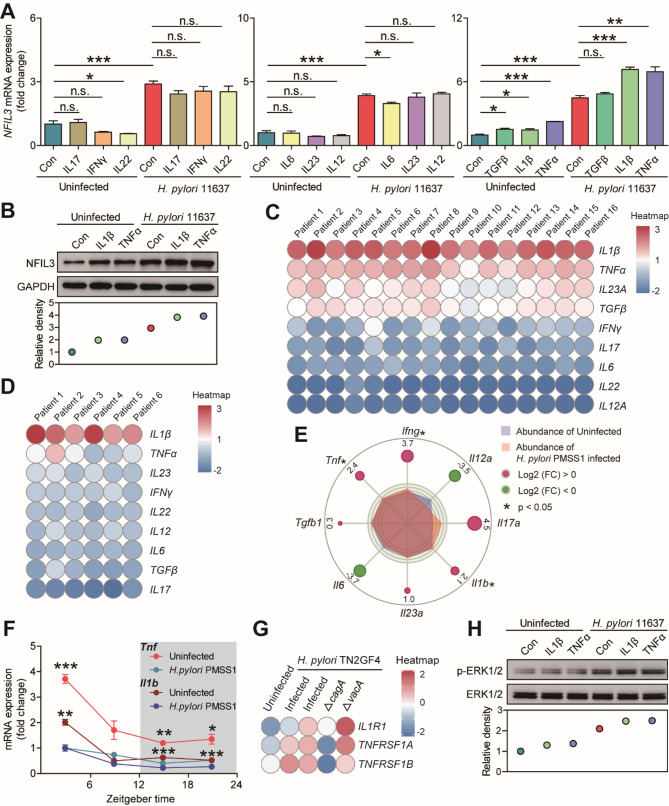
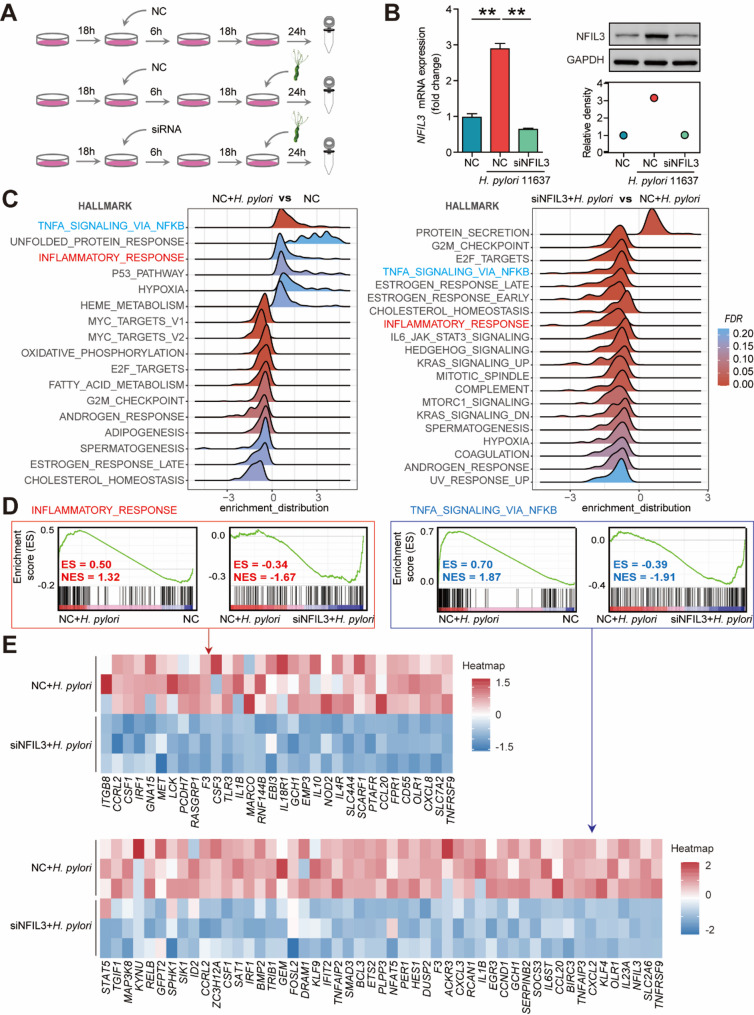
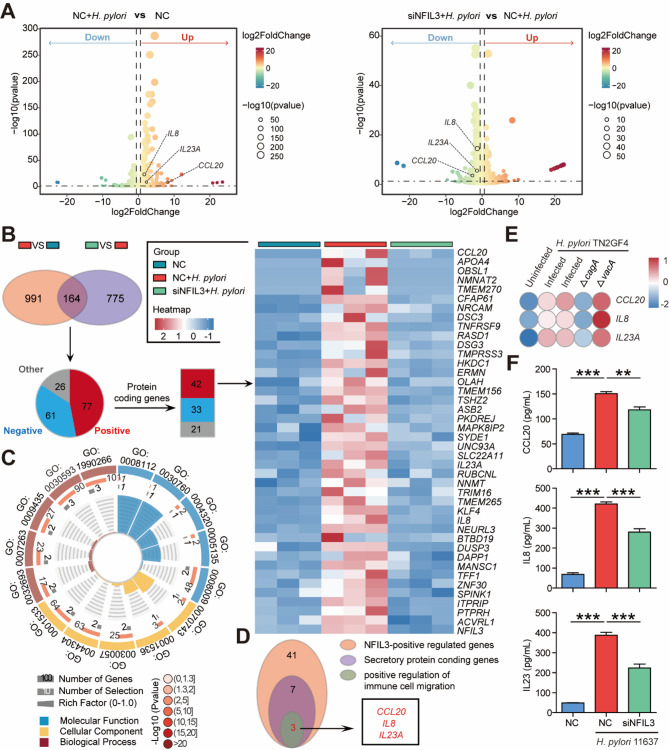
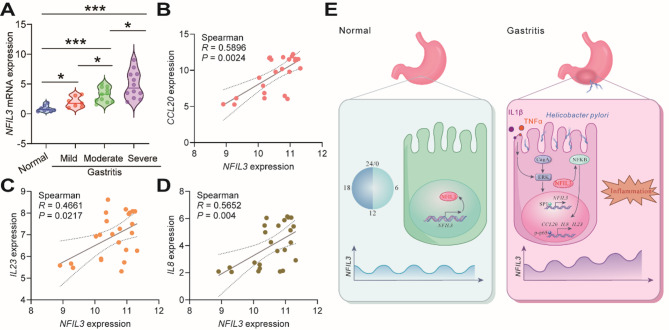
Helicobacter pylori (H. pylori) associated gastritis, marked by chronic gastric inflammation, heightens gastric cancer risk by fostering a malignancy-prone microenvironment. Disruption of the biorhythm contribute to the onset of various gastrointestinal disorders, such as gastric dyspepsia, gastric ulcers, and cancer. We aimed to investigate the functional roles and regulatory mechanisms of key biorhythm molecules in H. pylori associated gastritis. We investigated biorhythm gene expression in H. pylori-infected human gastric tissues and found significant impact on NFIL3 expression. Animal studies confirmed that H. pylori controls NFIL3 biorhythm. Clinical samples indicated a correlation between NFIL3 and gastritis severity, suggesting a regulatory role. Then, we found that H. pylori disrupt NFIL3 expression rhythm in gastric epithelial cells (GECs) through the CagA-activated ERK-SP1 pathway. Additionally, cytokines IL1β and TNFα enhance this disruption. RNA-seq and Gene set enrichment analysis (GSEA) indicated that NFIL3 positively regulates the inflammatory response during H. pylori infection. Our research highlights the crucial role of the biorhythm molecule NFIL3 in H. pylori associated gastritis. Modulating biorhythm molecules could be a promising therapeutic approach to manage disease progression, given their impact on gastrointestinal pathology.
Keywords: Helicobacter pylori; Biorhythm; Gastric epithelial cells; Gastritis; NFIL3.
© 2025. The Author(s).
Conflict of interest statement
Declarations. Ethics approval and consent to participate: This research was approved by the Ethical and Experimental Committee of The General Hospital of Western Theater Command and written informed consent was obtained from all patients before enrolling in the research program. Similarly, the animal experiments conducted have been approved by the Ethics Committee of the Ethical and Experimental Committee of The General Hospital of Western Theater Command. Consent for publication: All authors have read the manuscript and provided their consent for the submission. Competing interests: The authors declare no competing interests.
Figures
Similar articles
-
Expression of ETS1 in gastric epithelial cells positively regulate inflammatory response in Helicobacter pylori-associated gastritis.Cell Death Dis. 2020 Jul 1;11(7):498. doi: 10.1038/s41419-020-2705-8. Cell Death Dis. 2020. PMID: 32612120 Free PMC article.
-
Helicobacter pylori-induced IL-33 modulates mast cell responses, benefits bacterial growth, and contributes to gastritis.Cell Death Dis. 2018 May 1;9(5):457. doi: 10.1038/s41419-018-0493-1. Cell Death Dis. 2018. PMID: 29691371 Free PMC article.
-
Helicobacter pylori-induced REDD1 modulates Th17 cell responses that contribute to gastritis.Clin Sci (Lond). 2021 Nov 26;135(22):2541-2558. doi: 10.1042/CS20210753. Clin Sci (Lond). 2021. PMID: 34730176
-
Gastric epithelial cell proliferation and cagA status in Helicobacter pylori gastritis at different gastric sites.Scand J Gastroenterol. 2007 May;42(5):545-54. doi: 10.1080/00365520601014034. Scand J Gastroenterol. 2007. PMID: 17454874
-
Molecular response of gastric epithelial cells to Helicobacter pylori-induced cell damage.Cell Microbiol. 1999 Sep;1(2):93-9. doi: 10.1046/j.1462-5822.1999.00018.x. Cell Microbiol. 1999. PMID: 11207544 Review.
References
-
- Malfertheiner P, Megraud F, Rokkas T, Gisbert JP, Liou JM, Schulz C et al. Management of Helicobacter pylori infection: the Maastricht vi/florence consensus report. Gut. 2022.
-
- Li Y, Choi H, Leung K, Jiang F, Graham DY, Leung WK. Global prevalence of Helicobacter pylori infection between 1980 and 2022: a systematic review and meta-analysis. Lancet Gastroenterol Hepatol. 2023;8(6):553–64. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
